ГЛАВА 8 4 страница
Предположим, что в определенном сайте нормального гена (скажем, в 106-м положении) находится пара А-Т, а в том же сайте мутантного гена — G-C. Зная нуклеотидные последовательности, фланкирующие 106-й нуклеотид, можно синтезировать два коротких (20-нуклеотидных) фрагмента, прилегающих к данному сайту и комплементарных противоположным цепям (рис. 9.10). Основная особенность этой пары олигонуклеотидов состоит в том, что 3'-концевой нуклеотид одного из них (зонд X) комплементарен основанию, находящемуся в 106-м положении нормальной последовательности, а 5'-концевой нуклеотид второго (зонд Y) комплементарен нуклеотиду, примыкающему к 106-му нуклеотиду. При отжиге этих зондов с содержащей нормальную последовательность ДНК-мишенью (амплифицированной методом ПЦР) происходит их полная гибридизация, и при добавлении в реакционную смесь ДНК-лигазы зонды X и Y ковалентно сшиваются. Если же эти зонды отжигаются с мутантной ДНК, в которой произошла замена 106-го нуклеотида, то некомплементарный ему 3'-концевой нуклеотид зонда X не может образовать с ним пару. И хотя зонд Y по-прежнему гибридизуется полностью, ДНК-лигаза не может сшить зонды X и Y.
Молекулярная диагностика 197
| Рис. 9.10. Метод ПЦР/ЛОЗ. Б - биотин; Д — дитоксигенин; ЩФ - щелочная фосфатаза; СА — стрептавидин. | 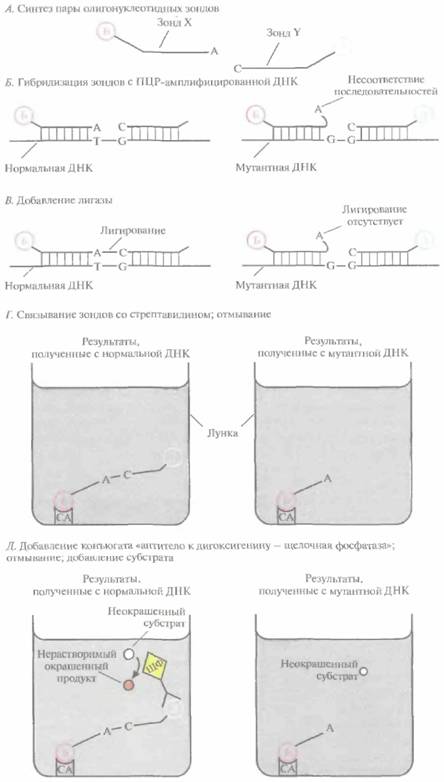
|
198 ГЛАВА 9
Можно синтезировать и другие олигонуклеотидные зонды, полностью соответствующие последовательности с мутантным 106-м нуклеотидом. При таком наборе зондов лигирование будет происходить в случае их отжига с мутантной ДНК-мишенью и не будет в случае отжига с нормальной мишенью. Таким образом, метод ПЦР/ЛОЗ различает две ситуации: лигирование зондов и отсутствие лигирования.
Чтобы определить, произошло ли лигирование, 5'-конец зонда X метят биотипом, а 3'-конец зонда Y — дигоксигенином, низкомолекулярным соединением, связывающимся с соответствующим антителом. После гибридизации и лигирования проводят денатурацию ДНК для высвобождения гибридизовавшегося зонда и переносят смесь в небольшую пластиковую лунку, покрытую стрептавидином. Лунку промывают, чтобы удалить весь материал, кроме связавшегося со стрептавидином биотинилированного зонда. Затем добавляют в лунку антитела к дигоксигенину, предварительно соединенные со щелочной фосфатазой. После промывания, в ходе которого происходит удаление несвязанного конъюгата, добавляют бесцветный хромогенный субстрат. Окрашивание раствора в лунке свидетельствует о связывании антитела к дигоксигенину с зондом, меченным дигоксигенином, т. е. о том, что этот зонд был лигирован с зондом, меченным биотином. Если же окрашивания не происходит, значит лигирования не было.
Располагая двумя парами зондов, можно установить генетический статус любого человека. Например, ДНК гетерозиготных носителей дает положительный ответ с обеими парами зондов, ДНК лиц, обладающих двумя копиями нормального гена, — только с тем набором зондов, который содержит нуклеотид, комплементарный нормальному сайту, и, наконец, ДНК индивидов с двумя измененными копиями гена — только с набором зондов, детектирующим мутантный сайт. Чтобы минимизировать необходимое для анализа количество исходной ДНК, перед гибридизацией участок ДНК-мишени, содержащий тестируемый сайт, амплифицируют с помощью ПЦР,
ПЦР/ЛОЗ является быстрым, чувствительным и высокоспецифичным методом. Все его стадии роботизированы, что позволяет проводить до 1200 тестов в день.
Более простым, хотя и менее чувствительным вариантом ПЦР/ЛОЗ является метод лигазной цепной реакции. Тестируемую ДНК смешивают с избытком двух индикаторных зондов, описанных выше, в присутствии термостабильной ДНК-лигазы. Проводят лигирование при 65 °С, затем повышают температуру до 94 °С, чтобы произошла денатурация образовавшихся гибридов зонд—ДНК-мишень, и вновь понижают температуру до 65 °С для гибридизации свободных нелигированных индикаторных ЛОЗ-зондов с ДНК-мишенью. Этот цикл повторяют 20 раз. Если индикаторные ЛОЗ-зонды полностью комплементарны ДНК-мишени, то лигирование будет происходить в каждом цикле, и после 20 циклов накопится достаточно продуктов лигирования («сшитых» зондов X и Y) для того, чтобы их можно было обнаружить с помощью электрофореза или ELISA. Если комплементарность неполная, то лигирование не произойдет и никаких продуктов зарегистрировано не будет.
Генотипирование с использованием флуоресцентно меченных ПЦР-праймеров
Колориметрическое генотипирование основано на применении ПЦР-праймеров, меченных различными флуоресцентными красителями. Чтобы различить мутантную ДНК и ДНК дикого типа, проводят ПЦР с двумя разными праймера-ми. Один из них (Р1) комплементарен ДНК дикого типа и на 5'-конце помечен родамином (красный цвет), другой (РЗ) комплементарен мутантной ДНК и на 5'-конце помечен флуо-ресиеином (зеленый цвет) (рис, 9.11). В обоих случаях амплификацию проводят в присутствии третьего, немеченного праймера (Р2), комплементарного противоположной цепи. Поскольку ПЦР может идти только в том случае, когда праймер полностью комплементарен ДНК-мишени, в присутствии в реакционной смеси всех трех праймеров будет амплифицироваться либо ДНК дикого типа, либо мутантная ДНК, либо обе они, в зависимости от ДНК-мишени, играющей роль матрицы. Если индивид гомозиготен по ДНК дикого типа, то после проведения ПЦР и удаления лишних праймеров будет наблюдаться флуоресценция красного цвета, если он гомозиготен по мутантной ДНК — зеленого, а если присутствуют и мутантная ДНК, и ДНК дикого
Молекулярная диагностика 199
типа (т. е. индивид гетерозиготен) — желтого. Этот метод можно автоматизировать и адаптировать для любого однонуклеотидного сайта-мишени в любом гене с известной нуклеотидной последовательностью.
Мутации в разных сайтах одного гена
Далеко не все генетические заболевания обусловливаются одним специфическим изменением в гене. В большинстве случаев мутации происхо-
| Рис. 9.11. Обнаружение точко- вой мутации с помощью флуоресцентно меченных ПЦР-праймеров. А. Используя праймеры Р1 и Р2, амплифицируют ДНК дикого типа. Мутантная ДНК при помощи данных праймеров не амплифицируется из-за несоответствия ей праймера Р1, 5'-конец праймера Р1 помечен родамином, праймер Р2 немеченый. Б. Используя праймеры РЗ и Р2, амплифицируют мутантную ДНК; ДНК дикого типа в этом случае не амплифицируется. 5'-конец праймера Р3 помечен флуорес ценном, праймер Р2 немеченый. Знаки «+*> и *— » соответствуют сайту дикого типа и мутантному сайту, В случае генотипов «+/+». «+/— » и «— /— » образуются ПЦР-продукты, содержащие только родамин, смесь родамина и флуоресцеина и только флуоресцеин, и соответсгвенно наблюдается Красная, желтая и зеленая флуорес ценция . | 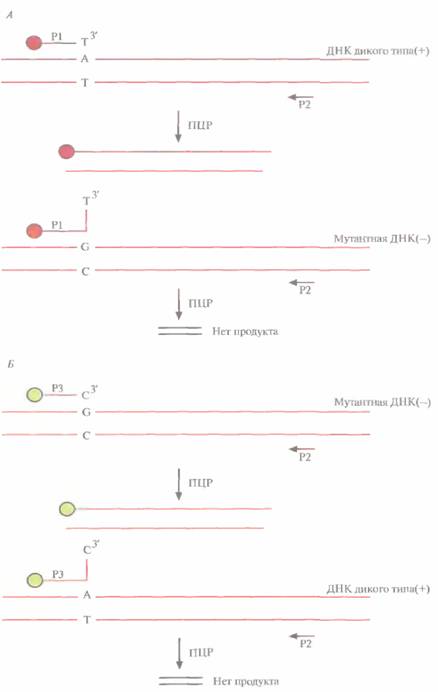
|
200 ГЛАВА 9
дят в разных сайтах в пределах одного гена, но приводят к одному генетическому заболеванию. В качестве примера можно привести ß-талассемию — наследственное заболевание, связанное с утратой активности ß-глобина. У гетерозиготных носителей при этом обычно наблюдается небольшая анемия. Индивиды же, гомозиготные по одному из как минимум восьми возможных мутантных сайтов, для поддержания жизни нуждаются в регулярном переливании крови и другом лечении. Поскольку мутация в любом из восьми специфических сайтов ß-глобинового гена может приводить к ß-талассемии, необходимо провести по крайней мере восемь разных тестов. Такая диагностика возможна, хотя и весьма дорогостояща.
Поэтому для скрининга мутаций, возникающих в разных сайтах одного гена, была разработана стратегия ПЦР/гибридизация, основанная на проведении одной реакции. Для этого синтезируют набор специфических 20-нуклеотидных зондов, каждый из которых полностью комплементарен фрагменту гена-мишени, несущему известную мутацию. К 3'-концу каждого зонда присоединен гомополимер poly(dT) длиной
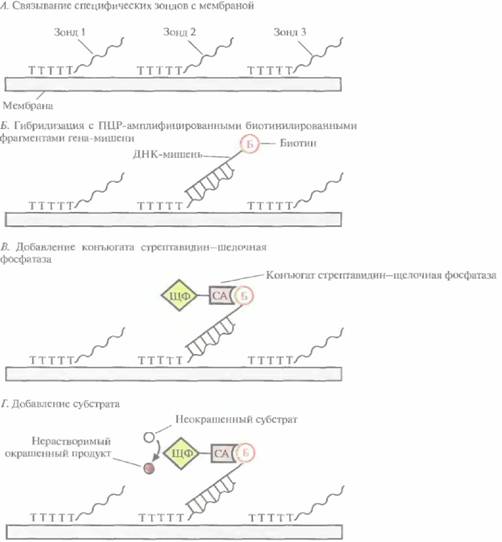
| Рис. 9.11. Выявление мутаций в разных сайтах одного гена. Б — биотин; СА -стрептавидин; ЩФ - щелочная фосфатаза. |
Микробиологическое производстволекарственных средств 201
примерно 400 нуклеотидов, с помощью которого ДНК-зонд связывается с заранее отмеченной точкой на найлоновом фильтре, а остальная его часть остается свободной и может гибридизоваться (рис. 9.12). Сегменты тестируемой ДНК, каждый из которых включает по одному из возможных мутационных сайтов, одновременно амплифицируют с помощью ПЦР, причем один пpaймер из каждой пары на 5'-конце помечен биотипом. Амплифицированные фрагменты ДНК-мишени гибридизуют с зондами, пришитыми к фильтру, в условиях, обеспечивающих гибридизацию только полностью комплементарных последовательностей. В гибридизационную смесь добавляют стрептавидин, связанный с щелочной фосфатазой (можно также использовать пероксидазу хрена или уреазу). После гибридизации промывают фильтр и добавляют неокрашенный субстрат. Если имеет место полное соответствие между амплифицированным сегментом ДНК-мишени и специфическим олигонуклеотидным зондом, то на фильтре появится цветная точка. На один и тот же фильтр можно нанести несколько точек, соответствующих целому ряду разных специфических олигонуклеотидных зондов. Проанализировав эту цветную мозаику, можно идентифицировать один из многих возможных сайтов мутации.
Перспективы
Молекулярная диагностика — это быстро развивающееся направление. Хотя его основные принципы уже сформировались, технические детали отдельных тестов могут различаться. Для получения в достаточном количестве ДНК-мишени сейчас успешно применяют ПЦР. Использование ПЦР и специфических зондов существенно повышает чувствительность тестов и позволяет применять нерадиоактивные хромогенные, хемилюминесцентные и флуоресцентные системы регистрации. Во многих случаях для выявления мутации или экзогенной ДНК инфекционного агента в исследуемом образце достаточно провести ПЦР с последующим электрофоретическим разделением продуктов. Не вызывает сомнения, что с помощью ДНК-диагностики можно будет выявлять большинство, а возможно и все наиболее распространенные генетические и инфекционные заболевания, а также новообразования.
ЗАКЛЮЧЕНИЕ
Любой эффективный диагностический тест должен быть: 1) высокоспецифичным в отношении молекулы-мишени; 2) достаточно чувствительным для выявления небольших количеств мишени; 3) достаточно простым, позволяющим без труда получать однозначные результаты. Существуют два типа методов молекулярной диагностики: один основан на сродстве антитела к конкретному антигену, другой — на идентификации специфических нуклеотидных последовательностей с помощью гибридизации или ПЦР.
Наиболее распространенным иммунологическим методом является ELISA. Вкратце он состоит в следующем: 1) фиксация образца на твердой подложке; 2) добавление первого антитела, специфичного к антигену-мишени, и его связывание с антигеном-мишенью; 3) добавление конъюгата второе антитело-фермент, который присоединяется к первому антителу; 4) добавление неокрашенного субстрата, который под действием фермента, входящего в состав конъюгата, превращается в окрашенное соединение. Изменение цвета реакционной смеси свидетельствует о присутствии в образце молекулы-мишени.
EL1SA применяется для обнаружения различных белков, идентификации вирусов и бактерий, а также определения низкомолекулярных соединений в широком спектре биологических образцов. Чтобы повысить специфичность первых антител, для диагностики часто используют моноклональные антитела. При этом для уменьшения стоимости прибегают к технике клонирования их фрагментов в Е. coli и получают комбинаторную библиотеку, а на ее основе — широкий спектр комбинаций Fv-фрагментов.
Высокочувствительным и специфичным методом обнаружения нуклеотидных последовательностей в биологических образцах является гибридизация. Его использовали при разработке способов идентификации патогенных микроорганизмов в клинических образцах и различных микроорганизмов в окружающей среде.
202 ГЛАВА 9
ДНК-диагностика основывается на обнаружении известных нуклеотидных последовательностей; для этого синтезируют специфические праймеры и амплифицируют последовательность-мишень. Это позволяет использовать нерадиоактивные системы детекции (например, хемилюминесцентный метод) или регистрировать ПЦР-продукты методом гель-электрофореза. Кроме того, ПЦР-продукты можно пометить флуоресцентным красителем, присоединив его к 5'-концу праймера.
В судебной медицине все более широкое применение находит метод геномной дактилоскопии, основанный на том, что ДНК каждого человека образует уникальный набор гибридизационных полос. При этом в качестве зондов обычно используют минисателлитные ДНК человека, которые не кодируют никаких белков и отличаются высокой вариабельностью.
Для характеристики ДНК растений используют набор произвольных олигонуклеотидных праймеров, проводят ПЦР-амплифицикацию случайных фрагментов ДНК, осуществляют электрофорез и получают специфичный для каждого растения набор полос ДНК; данный подход носит название RAPD.
Методы ДНК-диагностики применяют также для обнаружения точковых мутаций в данном гене. Один из подходов заключается в лигировании двух олигонуклеотидных праймеров. При несоответствии всего одного нуклеотида в месте стыковки гибридизовавшихся олигонуклеотидов лигирования не происходит.
ЛИТЕРАТУРА
Barany F.1991. Single-nucleotide genetic disease detection using cloned thermostable ligase. Proc. 1991 Miami Bio/Technol. Winter Symp. 1:88.
Barker R. H., L,Suebsaeng, W. Rooney, G.C. Alecrim, H. V.Dourado, D. F. With. 1986. Specific DNA probe for the diagnosis of Plasmodium falciparum malaria. Science 231: 1434-1436.
Bugawan T L., R. K. Saiki, С. H. Levenson, R.M. Watson, H.A. Erfich.1988. The use of non-radioactive oiïgonucleotide probes to analyze enzy-matically amplified DNA for prenatal diagnosis and forensic HIA typing. Bio/Technology 6:943-947.
Carlson D. Р., С. Superko, J. Mackey, M. E. Gaskill, P. Hansen.1990. Chemilumines-cent detection of nucleic acid hybridization. Focus 12:9-12,
Caskey C. T.1987. Disease diagnosis by recombinant DNA methods. Science 236: 1223-1229.
Chehab F. F., Y. W. Kan.1989. Detection of specific DNA sequences by fluorescence amplification: a color complementation assay, Proc. Natl. Acad. Set. USA 86: 9178-9182.
Debenham P. G.1992. Probing identity: the changing face of DNA finge [printing. Trends Biotechnol. 10:96-102.
Erlich H. A., D. Gelfand, J. J. Sninsky.1991. Recent advances in the polymerase chain reaction. Science 252: 1643-1651.
Gillani 1.C. 1987. Non-radioactive probes for specific DNA sequences. Trends Biotechnol. 5: 332-334.
Hartskeerl R.A., M. Y. L.. De Wit, P. R.Klatser. 1989. Polymerase chain reaction for the detection of Mycobacterium teprae. J. Gen. Microbiol, 135: 2357-2364.
Jeffreys A. J., A. MacLeod, K. Tamaki, D. L. Neil, D. G.Monckton. 1991. Mini satellite repeat coding as a digital approach to DNA typing. Nature 354: 204-209.
Kingsbury D.T. 1987. DNA probes in the diagnosis of genetic and infectious diseases. Trends Biotechnol. 5: 107-111.
Klevan L., G. Gebeyehu.1990. Biotinylated nucleotides for labeling and detecting DNA. Methods Enzymol. 184: 561-577.
Kohler G., С.Milstcin, 1975. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 256: 495—497.
Kuppuswamy M. N., J. VV. Hoffmann, C. K. Kasper, S. G. Spitzer, S. L· Groce, S. P. Bajaj.1991. Single nucleotide primer extensioin to detect genetic diseases: cAperimental application to hemophilia В (factor IX) and cystic fibrosis genes. Proc. Natl. Acad. Sei. USA 88:1143-1147.
Mathews J. A., L. J. Kricka.1988. Analytical strategies for the use of DNA probes. Anal. Biochem. 169: 1-25.
Nickerson D. A., R. Kaiser, S. Lappin, J. Stewart, L Hood, U. Landegren.1990. Automated DNA diagnostic using an ELlSA-based oligonucleotide ligation assay. Proc. Natl. Acad. Sei. USA 87: 8923-8927. '
Микробиологическоепроизводство лекарственных средств 203
Дата добавления: 2015-07-14; просмотров: 1132;