Программирование поведения
Одним из основных атрибутов любого сложного целенаправленного движе-ни является формирование предварительных программ.
Роль программы в структуре двигательного акта должна рассматриваться с учетом биологической мотивированности движения, его временных параметров, моторной дифференцированнос-ти, степени сложности координационного состава и уровня его автоматизи-рованности.
Стратегия и тактика движения. Биологическая мотивированность двигательного акта является основным побуждающим (инициальным) фактором для его реализации. Именно мотивации формируют целенаправленные движения, а следовательно, определяют их общую стратегию. Значит, если в основе стратегии движения находится биологическая (или социальная) мотивация, то каждый конкретный двигательный акт будет рассматриваться как шаг к удовлетворению этой мотивации, т. е. будет решать какую-то промежуточную задачу или цель (рис. 72). Биологические мотивации могут приводить либо к запуску «запаянных», т. е. жестких, программ, устанавливать их комбинаторику, с чем мы встречаемся у беспозвоночных и низших позвоночных и именуем инстинктами или комплексами фиксированных действий, либо приводить к формированию новых сложных программ, определяя одновременно степень их лабильности.
П. Тейтельбаум (1967) пишет, что в тех случаях, когда действие полностью является автоматическим следствием стимула, невозможно говорить о моти-
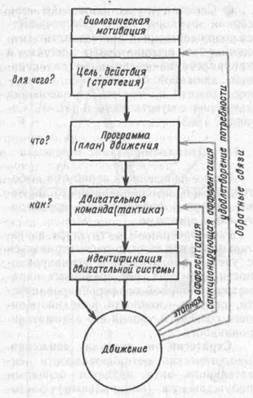
Рис. 72- Соотношение прямых и обратных связей в системеуправления движением
вации. При этом между стимулом и ответом существуют фиксированные взаимоотношения. Мотивация «ломает» эти фиксированные связи между стимулом и реакцией с помощью процесса обучения. Например, в отличие от многих инстинктивных реакций реакция нажатия на педаль может быть «отделена» от внутреннего состояния животного. Оперантная ситуация, сигнал, реакция, подкрепление являются полностью произвольными, не обладающими фиксированными связями друг с другом. Возьмем для примера такое биологическое адекватное движение у кошек, как прицельный прыжок. У одной группы кошек он условно-рефлекторно закреплялся при пищевом подкреплении, у другой — при электрооборонительном (А. С. Батуев, 1960). У животных экстренно выключалась зрительная рецепция. Условные прыжки при пищевом подкреплении исчезли полностью.
а при электрооборонительном — восстанавливались через две недели после операции. Следовательно, одна и та же двигательная программа (или план движения, по Прибраму) характеризовалась разной степенью устойчивости к сенсорной депривации в зависимости от биологического знака подкрепляющего агента. Образно говоря, если мотивация определяет, для чего, с какой целью осуществляется движение, то она же, определяя программу конкретного движения, обусловливает степень ее зависимости от сенсорных факторов. В качестве обратной афферентации здесь выступает удовлетворение мотивации. Поэтому цель движения, его стратегия должна быть прежде всего адаптивна по отношению к доминирующей биологической мотивации.
Программа определяет, какое конкретное движение будет осуществляться для достижения данной цели, и формирует его пространственно временные параметры в соответствии с реальной обстановкой. Следовательно, программа движения должна быть в высокой степени адаптивной ко всем факторам окружающей среды и строится при их одновременном и строгом учете, т. е. на основе синтеза всех видов афферентации по закону мультисенсорной конвергенции.
В общей схеме системной организации двигательного акта существует понятие о тактике движения, которая определяет, как будет осуществляться требуемое движение, иными словами, каковы те эфферентные залпы к двигательным ядрам спинного мозга, которые определяют активацию различных мышечных групп. Последовательность включения той или иной конечности, записанная в программе, реализуется в виде последовательности импульсных залпов надсегментарных структур к различным мотонейронным пулам.
В отличие от программы двигательные команды должны быть очень адаптивными к функциональному состоянию самого скелетно-двигательного аппарата как непосредственного исполнителя этих команд. Исходя из представления о существовании настройки перед со-
вершением движения (Я. М. Коц, 1975; Э. Эвартс, 1975), высказано предположение о том, что срочная идентификация состояния сложной системы осуществляется путем оценки реакции системы на небольшое точно дозированное возмущение (тест-импульс), которое подается непосредственно перед или с моментом запуска команд. Причем тест-импульс подается только на те мышцы, которые должны будут участвовать в следующем движении. Реакция мышц на тест-импульс будет не настолько значительна, чтобы существенно изменить само движение, но достаточна для возбуждения мышечных веретен при изменении длины интрафузальных волокон па 5—10 мк. Информация о реакции на тест-импульс конвергирует на некотором промежуточном уровне с основным управляющим воздействием (командой), которое корректируется в соответствии с актуальным состоянием двигательного аппарата. Учитывая временную дисперсию проведения по пирамидному тракту, легче всего допустить участие быстропроводящих его волокон (со скоростью 50—60 м/с) в реализации такого тестового воздействия и коррекции последующих нисходящих импульсных залпов уже на сегментарном уровне. Таким образом, биологически мотивированная стратегия (цель) движения, соответствующая программа конкретного двигательного акта, иерархически построенные центральные команды и система идентификации состояния мышечного аппарата представляют собой важнейшие звенья организации двигательного акта, каждое из которых корректируется соответствующими обратными связамн.
Последние обеспечивают движению высокую адаптивность в конкретных условиях времени и пространства.
Участие ассоциативных систем мозга в организации движений. Роль внешних факторов, сигналов из внешней среды и, соответственно, роль сенсорных и ассоциативных систем мозга в формировании мотивированных движений весьма значительна. Специфичность участия таламопариеталыюй ассоциативной системы в организации движений определяется двумя моментами: 1) с одной стороны, она участвует в формировании интегральной схемы те-
ла, все части которого соотнесены не только друг с другом, но и с вестибулярными и зрительными сигналами; 2) с другой стороны, она участвует в регуляции внимания к текущим сигналам окружающей среды с учетом ориентации всего тела относительно этих сигналов. Таламопариетальная (как и нижневисочная) ассоциативная система активируется текущими сенсорными сигналами, т. е. привязана в основном к настоящему моменту времени, и связана с анализом главным образом пространственных взаимоотношений разномодальных признаков.
Фронтальная ассоциативная система имеет рсципрокные отношения с двумя функциональными системами мозга: 1) теменно-височной, которая связана с обработкой и интеграцией полимо-далышй сенсорной информации, и 2) те-ленцефалический лимбической системой, включающей лимбическую кору и связанные с ней подкорковые образования, особенно гипоталамус и районы среднего и промежуточного мозга. Характер связей указывает на то, что фронтальная система вовлечена и в сенсорные, и в моторные механизмы мозга (рис. 73). Вентральную часть фронтальной коры рассматривают как кортикальную проекцию лимбической системы, которая участвует в анализе и регуляции процессов внутренней среды организма.
Одно из существенных функциональных расстройств, возникших у человека после поражения лобных долей мозга, это распад поведенческих программ, неспособность поддерживать нормальную временную организацию поведения. ' Программы их действий, будучи однажды запущенными, повторяются, застаиваются и лишь постепенно стираются, причем эта персеверация зачастую приводит к отклонению от желаемой цели (рис. 74). Тот факт, что даже осознаваемое различие между целью и результатом не ведет к смене программы, предполагает, что все эти сигналы — ошибки, обычно корректирующие поведенческие программы, не находят выхода в эффекториую сферу. Отсюда следует, что фронтальная

Рис. 73. Схема взаимодействия различных структур мозга, ответственных заформирование етпатегии и тактики двигательного поведения (по X. Корнхуберу. 1974):
Дата добавления: 2015-08-04; просмотров: 1486;