AM — передние медиальные ядра таламуса; VL — вентролатеральное ядро таламуса; ГП — генератор плавных движений; ГС - генератор движений саккадического типа
ассоциативная кора опосредует мотива-ционные влияния на организацию поведения в целом.
Реципрокные фронто-лимбические взаимоотношения лежат в основе феномена «поведенческого предвосхищения», или экстраполяции, потеря которой является одним из важных клинических симптомов фронтальных повреждений. Обычно нормальный субъект выбирает линию поведения после сравнения большего или меньшего числа альтернативных тактик. В конечном счете этот выбор осуществляется при сравнении . эффективных реакций на каждую из возможных альтернатив. Выдвинутая таким образом тактика естественно должна пройти «цензуру» мотивационной системы. Процессы такого предвосхищающего (экстраполя-ционного) выбора существенно страдают при удалении лобной коры.
Утрата фронтальной коры как главного передатчика информации между
корой и лимбической системой будет сопровождаться не только нарушением выбора программ, но также стиранием текущей или предполагаемой программы действия самой по себе или в результате вмешательства внешних сигналов. Нарушение процессов кратковременной памяти при лобном синдроме может быть вызвано патологически быстрым угасанием следов сигналов, с одной стороны, и преобладанием «теменных» механизмов переключения внимания на новые стимулы, освобожденные от тормозных фронтальных влияний, с другой стороны (Б. Мильнер, 1964, 1975).
Импульсные коды моторных программ.Программирующая деятельность мозга заключается в организации поведенческого акта на основе доминирующей мотивации, прошлого жизненного опыта и в соответствии с вероятностной структурой среды. Степень адекватности программы, построенной с учетом всех трех факторов,
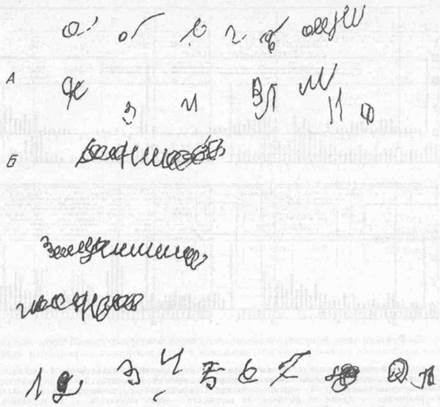
Рис. 74. Примеры почерка больного с локальным поражением правой лобной доли.
[Видны затруднения в написании цифр и букв (А), а также навязчивые повторные движения (персеверации) при попытке написания слов (Б)}
может быть оценена по ее прогнозирующим свойствам. В программе тесно взаимодействуют процессы кратковременной и долговременной памяти.
Для выявления импульсного кода моторных программ ситуация отсроченного пространственного выбора оказалась весьма удобной (А. С. Батуев и др., 1988). В опытах на обезьянах вырабатывались пищедобывательные условные рефлексы, сигналами для которых служило вспыхивание лампы справа или слева от животного, а условной реакцией — нажатие рукой, соответственно, на правый или левый рычаг. Такая деятельность животного основывается на долговременной памяти. Кроме того, в задаче фигурировали два вероятностно варьируемых фактора: чередования пространственного поло-
жения условного сигнала и продолжительность следуемой за ним временной паузы — отсрочки.. После восприятия положения сигнала животному предстояло сохранить полученную информацию на протяжении всего периода отсрочки — кратковременная память. Степень адекватности программирования оценивали по правильности выбора рычага, после нажатия на который следовало пищевое подкрепление. Таким образом, у животного формировались две жесткие моторные программы, которые извлекались из долговременной памяти и удерживались в кратковременной памяти вплоть до их реализации. Какая программа извлекалась — это зависело от пространственных признаков условного сигнала и определялось по моторному ответу. Следовательно,
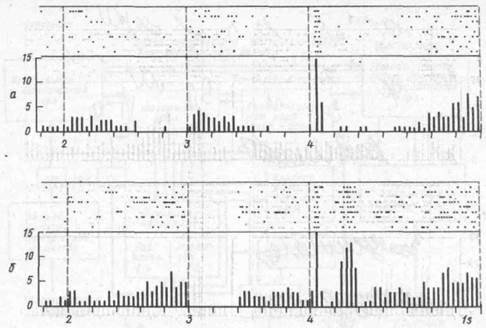
Рис. 75. Реакции одного пространственно селективного нейрона лобной коры обезьян но время выполнения целенаправленного движения с альтернативным отсроченным выбором:
а — растры и соответствующие гистограммы при действии условного сигнала справа; б — то же, слева; 2 3 период действия условного сигнала (загорание лампы после рычага); 3—4 — период отсрочки (кратковременной памяти); 4 — пусковой сигнал (открывание экрана, позволяющее осуществить манипулирование с одним из рычагов, по вертикали число импульсов, с, по горизонтали время, с, А t = 200 мс, n=10)
характер импульсной активности нейронов от начала действия условного сигнала, на всем протяжении отсрочки, после открытия доступа к рычагам (пусковой сигнал) и вплоть до нажатия на рычаг может рассматриваться как импульсный код моторных программ. '"'"•"'
Нейроны лобной и задней теменной
коры характеризовались пространственной селективностью, т. е. кодированием местоположения условного сигнала рисунком или частотой разряда (рис. 75). Такие же клетки найдены в хвостатом ядре и вентральном переднем ядре таламуса, но они отсутствовали в бледном шаре, моторной коре, медиодорзальном и центральном латеральном ядрах таламуса. Одна часть клеток обнаружила различия в активности только в момент действия условных сигналов («сенсорные нейроны») или только в период отсрочки («нейроны памяти»). Как отмечалось, адекватность извлеченной программы можно оценить только по конечному результату.
В числе пространственно селективных клеток обнаружена популяция нейронов, которые выявляют это свойство только после открытия экрана, т. с. в пусковой период, когда животное получает свободный доступ к рычагам. Их назвали нейронами «моторных про грамм», которые эстафетно активируются нейронами памяти и являются пусковыми для соответствующего двигательного акта (рис. 76). Отчетливо видно, что в случаях правильного выбора рычага при одном и том же движении, но направленном либо к правому (сверху), либо к левому (снизу) рычагу, возникает совершенно противоположный по характеру рисунок импульсной активности нейрона. То же наблюдается и при ошибочном выборе рычага для нажатия.
Гистограмма правильного пыбора рычага на правый сигнал отражает активацию моторной программы нажатия на правый рычаг. Но при ошибоч-
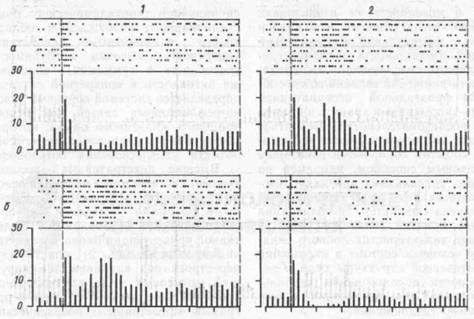
Рис.76. Активность нейрона лобной коры обезьяны послепускового сигнала(открывание экрана)
а — пространственный условный сигнал был дан справа; б — слева; I — правильный выбор рычага; 2 — ошибочный выбор; стрелка - момент открывания экрана. Видно, что рисунок гистограммы соответствует планируемому и выполняемому движению с правым {1а и 26) или с левым рычагом (16 и 2а) "
ном выборе по условному сигналу слева животное вновь нажимает на правый рычаг. То же справедливо и в случаях нажатия на левый рычаг. Сравнение гистограмм по диагонали убеждает в их полной идентичности. Значит, рисунок разряда нейрона при нажатии на правый или левый рычаг отражает лишь специфику извлеченных ранее из долговременной памяти жестких моторных программ. Рисунок разряда нейрона отражает основное различие в этих программах — направление двигательного акта. Обнаружены также смешанные типы нейронных популяций, которые описывают местоположение условного сигнала, сохраняя эту особенность импульсации в период отсрочки и до реализации соответствующего двигательного акта. Такие нейроны характерны для лобной, теменной коры, хвостатого ядра и вентрального переднего ядра. Другие исследованные структуры (моторная кора, бледный шар и др.), вероятно, не имеют отношения к фор-
мированию моторных программ, но тесно связаны с реализацией соответствующего движения.
• Анализ импульной активности нейронов позволил допустить существование достаточно жестких форм программирования движений, при которых вероятностный сигнал сразу встраивается в уже готовую конструкцию сложного сенсомоторного синтеза, выработанного в процессе предварительной условно-рефлекторной тренировки. Выбор двух возможных действий выступает как извлечение из долговременной памяти адекватной моторной программы с одновременным торможением программы альтернативной. Торможение выступает в качестве ведущего процесса ограничения степеней свободы для вывода на поведенческий уровень единственно адекватной двигательной программы.
Таким образом, самой сложной функцией мозга является способность формировать последовательность дви-
жений и предвидеть ее реализацию. Морфологически решение этой задачи связано с фронтальной ассоциативной системой. Возможность воспроизведения запрограммированной последовательности движений связана со способностью фронтальной ассоциативной системы запоминать и хранить в памяти такие последовательности. Вероятнее всего эти последовательности и содержание событий кодируются некоторым сокращенным способом, например по ведущему сигнальному признаку. Высшим отражением этого кодирования у человека является вербализация основных понятий движения. Одна из основных характеристик лобного синдрома у человека состоит в нарушении грамматической структуры речи и невозможности использования вербальных команд для построения последовательности движений.
* * *
Целенаправленное поведение определяется доминирующей мотивацией, побуждающей организм к удовлетворению преобладающей потребности. Адаптивный же характер поведения достигается с помощью множества условных рефлексов, которые обеспечивают приспособление организма к конкретной пространственно-временной ситуации. Неспецифическая направленность поискового поведения определяет-
ся наличием гипоталамического очага стационарного возбуждения, обладающего доминантными свойствами (инертность, высокая возбудимость, способность к суммации); поисковая же активность в конкретной ситуации определяется системой корковых условно-рефлекторных связей как основы прошлого жизненного опыта, который обеспечивает направленный поиск объекта удовлетворения потребности.
Высшие интегративные (ассоциативные) системы мозга являются основными аппаратами управления пластичными формами поведения, которые обеспечиваются механизмами 1) селективной конвергенции биологически значимой информации, 2) пластических перестроек под влиянием доминирующей мотивации и 3) краткосрочного хранения интегральных образов и программы предстоящего поведенческого акта. Степень развития ассоциативных систем мозга в эволюции млекопитающих коррелируют с совершенством аналитико-синтетической деятельности и организацией сложных форм поведения.
Способность формировать последовательность движений и предвидеть ее реализацию как самая сложная функция мозга достигает наибольшего развития у человека, обладающего свойствами речевого управления поведением.
Глава
Дата добавления: 2015-08-04; просмотров: 1989;