Высшие интегративныесистемы мозга
Ретикуло-стволовый уровень интеграции
Ретикулярная формация — это филогенетически старая система мозга, которая не представляет собой единого анатомического целого и морфологически является гетерогенным образованием. Трудно определить и анатомическую протяженность ретикулярной формации, которая занимает центральное положение на всем протяжении ствола (рис. 68).
Нейрональной основой интегрирующей функции ретикулярной формации служат длинноотросчатые клетки, аксоны которых могут распространять влияние как в каудальном, так и в ростральном направлении. Ретикулярные клетки соединяют разные отделы внутри ретикулярной формации, а последнюю со спинным мозгом, большими полушариями и мозжечком.
Каждая сенсорная система направляет пути в ретикулярную формацию. Описаны мощные влияния соматических и висцеральных нервов, что позволило считать одной из функций рети-
кулярной формации висцеро-соматичес-кую интеграцию. Показаны пути в ретикулярную формацию от ядер тройничного нерва, вестибулярных ядер, верхней оливы.
Кора мозга оказывает регулирующие влияния на деятельность ретикулярной формации. Пути из ретикулярной формации в нисходящем направлении — к спинному мозгу, мозжечку, ядрам специфических систем и в восходящем — к структурам больших полушарий вплоть до коры также построены по топическому принципу, что свидетельствует о функциональной специализации ретикулярных ядер.
Для активности ретикулярных нейронов характерна аутогенная ритмика, которая служит для осуществления тонических влияний на спинной мозг и большие полушария головного мозга. Сами ретикулярные нейроны способны отвечать на раздражения практически всех афферентных путей или центральных мозговых структур. Такая обширная конвергенция влияний у разных ретикулярных единиц значительно варьирует и зависит от функционального состояния мозга. Если ретикулярный нейрон дифференцирует входные влияния по их модальностям, то полисинап-
тические цепи нейронов с короткими аксонами, по которым активность распространяется внутри самой ретикулярной формации, лишают первоначальный сигнал его модальной специфичности. Поэтому к ядрам таламуса поступает активация, являющаяся продуктом полимодального интегрирования на ретикулярных нейронах.
Дифференцированный характер конвергенции, топическое фракционирование и гетерогенность в организации афферентных и эфферентных связей позволяют заключить, что функции, приписываемые ретикулярной формации, касаются главным образом интеграции и регуляции разных видов деятельности.
П. К. Анохин (1968) обращал внимание на гетерохимичсскую чувствительность отдельных синапсов и целых полинейрональных цепей, активируемых раздражениями различного биологического качества (боль, голод и др.)-
Ретикулярные влияния по соответствующим путлм могут достигать различных релейных ядер сенсорных систем и дифференцированно модулировать афферентные потоки. Показано, например, что концентрация внимания животного на зрительном объекте приводит к блокированию передачи сенсорных импульсов в слуховой системе (П. Эр-нандец-Пеон, 1962). Явления, именуемые привыканием, связывают с регулирующей функцией ретикулярной формации. Поскольку привыкание рассматривают как простейший вид обучения, то ретикулярной формации приписывают известную роль в формировании условных рефлексов (А. Гасто, И. Иошчи и др.).
Интегрированные на ретикулярном уровне размодальные афферентные посылки с учетом мотивационных факторов формируют импульсные тонические влияния к специфическим сенсорным системам, приводящие к модуляции проведения по ним соответствующих сигнализаций. В результате создаются предпосылки для успешного синтеза различных сенсорных сигнализаций на более высоких уровнях мозга.
Другая сторона восходящих ретикулярных влияний может заключаться в создании на таламокортикальном уровне такого функционального состояния {или центрального тонуса мозга), который обеспечит адекватные условия для формирования разномодального афферентного синтеза.
Таламокортикальный уровень интеграции.
Нейрофизиология ассоциативных систем мозга.Наряду со специфическими и неспецифическими системами принято выделять в качестве самостоятельной категории ассоциативные таламокортикальные системы. Применительно к высшим млекопитающим это своеобразные структуры, не принадлежащие какой-либо одной сенсорной системе, но получающие информацию от нескольких сенсорных систем. Ассоциативные ядра таламуса относятся к «внутренним ядрам», афферентные входы к которым идут не от сенсорных лемнисковых путей, а от их релейных образований. В свою очередь, эти ядра проецируются на ограниченные корковые территории, которые именуются ассоциативными полями.
Согласно анатомическим данным выделяют две высшие ассоциативные системы мозга. Первая включает заднюю группу ассоциативных ядер, проецирующихся на теменную область коры, и именуется таламопариетальной системой. Вторая на таламическом уровне состоит из медиодорзального ядра с его проекцией на лобную область коры и называется таламофронтальной системой. Обе ассоциативные системы — продукт прогрессивной дифференциации неспецифического таламуса и достигают значительных размеров у приматов и человека.
Таламопариетальная система.Теменная кора является местом широкой гетеросенсорной конвергенции по волокнам от специфических, ассоциативных и неспецифических ядер таламуса, а также по путям от сенсорных корковых зон и симметричной коры противоположного полушария.
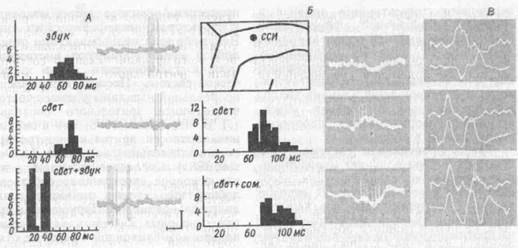
Рис. 69. Типы взаимодействия разномодальных сигналов на нейронах теменной коры мозга кошек
на А и Б: слева — постстимульные. гистограммы {по оси абсцисс длительность латентного периода, мс; по оси ординат -- число потенциалов действия); справа — импульсная активность нейронов; В суммарная (снизу) и импульсная (сверху) активность, сверху вниз на А: ответ на звук, свет и звук 4~t-'Bt-"r- на £ и В — ответ на соматическое раздражение; свет и свет + соматическое раздражение {два разных опыта). Калибровка: 250 мкВ, 100 мс. На схеме: ССИ -- средняя супрасильвиева извилина, точка — место расположения микроэлектродных треков
Световые воздействия оказывают наиболее сильный эффект на теменную кору: здесь описаны наряду с мульти-сенсорными и собственно зрительные нейроны, которые отвечают на перемещения в определенном направлении сложных геометрических фигур. Иногда эти же клетки отвечают и на звуковые воздействия, и на движения глаз.
Ф Обнаружено три типа межсенсорного взаимодействия на нейронах теменной коры: I — суммирующий нейрон, который при одновременном гете-росенсорном раздражении отвечает более сильным разрядом с более коротким латентным периодом, чем на мономодальные стимулы; II тормозящийся нейрон, латентный период ответа которого увеличивается, когда стимул из моносенсорного превращается в ге-теросенсорный и III — детектор комплекса, который отвечает импульсным разрядом только при комплексном ге-теросенсорном воздействии и не реагирует на изолированное применение мономодальных стимулов (рис. 69). Показано при этом, что в теменной коре существуют полисенсорные клетки, отражающие докорковый, таламический уровень интеграции, наряду с такими
нейронами, на мембране которых реализуются собственно кортикальные механизмы межсенсорного синтеза.
Теменная ассоциативная кора посылает мощные нисходящие связи ко многим сенсорным, лимбическим, ретикулярным и двигательным аппаратам мозга и даже образует волокна в составе кортикоспинального тракта (О. С. Адрианов, 1976).
Все вышесказанное дает основание рассматривать теменные области коры как важнейший дискриминационно-интегративный аппарат больших полушарий.
Ф После удаления теменных полей коры возникают глубокие нарушения в условно-рефлекторной деятельности как на простые мономодальные, так в особенности на разномодальные комплексные сигналы. Это связано со специальной ролью теменной коры в управлении процессами селективного внимания, с оптимазацией режима текущей деятельности, а также с формированием ориентационных движений к опознаваемому сигналу.
В целом таламопариетальная ассоциативная система мозга представляет собой: 1) центральный аппарат
первичного симультанного анализа и синтеза обстановочной афферентации и запуска механизмов ориентационных движений, 2) один из центральных аппаратов «схема тела» и сенсорного контроля текущей двигательной активности и 3) важнейший элемент долоб-ной предпусковой интеграции, участвующий в формировании целостных полимодальных образов.
Таламофронтальная система.Сен-сомоторная кора получаст множественные проекции из специфических, ассоциативных и неспецифических отделов таламуса, ассоциативные кортико-кор-тикальные и транскаллозальные входы из противоположного полушария и характеризуется наличием сложных си-наптических комплексов для конвергенции на одном и том же нейроне множества афферентных влияний. Не только вставочные нейроны, но и выходные элементы сенсомоторной коры — нейроны пирамидного тракта — относятся к категории мультисенсорных клеток.
ф Установлено три типа ответных реакций мультисенсорных нейронов сенсомоторной коры: 1) привыкание, заключающееся в снижении вероятности ответа на каждый последующий стимул в серии; 2) сенситизация — усиление реакции при повторных применениях стимула и 3) экстраполяция— формирование упреждающего ответа нейрона на каждый последующий стимул в серии.
Такие пластические перестройки ответной активности нейронов в зависимости от характера стимуляции и функционального состояния, интенсивности и модального состава стимулов свидетельствуют о наличии сложных механизмов функциональной конвергенции, имеющих непосредственное отношение к формированию системных реакций целостного организма.
Этого нельзя сказать по отношению к собственно лобным полям на дор-золатеральной поверхности прореаль-ной извилины кошек, которые по нейрофизиологическим и морфологическим данным еще не имеют непосредственной увязки с интегративными
процессами мозга, а скорее относятся к структурам неспецифического типа. В то же время мсдиобазальные отделы лобной коры формируют нисходящие пути к таламическим ядрам лимби-ческой системы. Последствия удаления прореальной извилины у кошек состоят в дефиците зрительного опознания (Л, В. Черенкова, 1975) или в нарушениях точности зрительно контролируемых двигательных актов (Ю. А. Юна-тов, 1981). Последнее связывают с тем, что основное пространство собственно лобной коры кошек занимает глазодвигательное поле 8. Нарушений в других сенсорных системах или эмоцио-нально-мотивационной сферы не отмечалось.
0 Таким образом, можно выделить следующие основные механизмы работы ассоциативных систем мозга (А. С. Ба-туев, 1979, 1981, 1984).
1. Механизм .мультисенсорной конвергенции. Его специфичность определяется тем, что к ассоциативным полям коры конвергируют афферентные посылки, несущие информацию о биологической значимости того или иного сигнала. Такие отселектированные афферентные влияния вступают в интеграцию на кортикальном уровне для формирования программы целенаправленного поведенческого акта.
2. Механизм пластических перестроек при гетеромодальных сенсорных воздействиях. Динамический характер мультисенсорной конвергенции может проявляться либо в избирательном привыкании, либо в сенситизации, либо наконец, в формировании экстраполя-ционного типа ответов. Установлен факт важной роли доминирующей мотивации в определении спектра конвергирующих модальностей и в организации внутрикорковых интеграции. Для теменной коры преобладающим может быть горизонтальный (ламинарный) тип межнейронной интеграции, а для сенсомоторной ассоциативной коры преобладающим является вертикальный (модульный) тип межнейронной интеграции.
3. Механизм краткосрочного хранения следов интеграции, заключающийся
в длительной внутрикорковой или тала-мокорковой реверберации импульсных потоков (см. гл. 6). Последнее объясняет дефекты памяти и обучения у кошек и собак после разрушения ассоциативных полей коры или соответствующих ядер таламуса.
Эволюция ассоциативных систем
В параллельных рядах, которыми шло развитие современных млекопитающих, хотя и сохранился общий план конструкции мозга, но таламо-кортикальные его системы претерпели наиболее существенные морфофункцио-нальные перестройки. Высокого развития достигают корковые механизмы деятельности сенсорных систем с ясно выраженной тенденцией возрастания ассоциативных систем мозга со свойствами полисенсорного конвертирования. В динамике морфологических преобразований происходит обособление зон перекрытия корковых проекций, с которыми связывают реализацию наиболее сложных форм высшей нервной деятельности.
В пределах класса млекопитающих можно выделить три основных уровня эволюции ассоциативных систем мозга. При этом необходимо иметь в виду, что степень развития ассоциативных формаций мозга рассматривается как показатель филогенетического статуса вида (Г. И. Поляков, 1964). Причем мозг насекомоядных рассматривается как предшественник с его дальнейшим усложнением в параллельных рядах грызунов, хищных и приматов. Исследования обнаружили нечеткость границ дифференциации внутри новой коры и неопределенность функциональной идентификации ее полей. Это согласуется с отсутствием в та ламу се четких границ между сенсорными ядрами.
Вместе с этим определенная область коры ежей (рис. 70) по морфологическим критериям позволяет допустить присущие ей интегративные свойства даже в пределах столь примитивно организованного неокортекса (Г. П. Демьяненко, 1977). К этой области коры ежей проецируются примитив-
ные ассоциативные ядра таламуса— медиодорзальное и заднелатеральное. В данной области обнаружены как разномодальные моносенсорные, так и полисенсорные нейроны (А. А. Пирогов, 1977). Такие корковые элементы активируются преимущественно по единому каналу, берущему начало в недифференцированной задней группе ядер таламуса. Таким образом в этой тала-мокортикальной системе сочетаются свойства как неспецифических, так и ассоциативных систем млекопитающих (рис. 70).
Было показано (И. В. Малюкова, 1974), что такая форма зрительно-слуховой интеграции, как двигательный условный рефлекс на одновременный комплекс, не может быть выработана у ежей и попытка ее сформирования приводит к невротическим срывам. Однако сложный цепной двигательный условный рефлекс может быть легко образован на мономодальный сигнал и разрушается после удаления ассоциативной области неокортекса. Больше страдают самые тонкие компоненты сенсомоторной интеграции, которыми завершается пишедобывательный двигательный акт.
Следовательно, хотя примитивная ассоциативная система ежей еще не способна к организации сложных актов межсенсорной интеграции, но она уже начинает участвовать в осуществлении процессов сенсомоторного синтеза.
Как уже указывалось, насекомоядные являются прямыми филогенетическими предшественниками грызунов, хищных и приматов.
Исследование конструкции ассоциативных систем мозга у крыс, кроликов, кошек, собак и низших обезьян позволили прийти к следующему заключению.
Первый уровень — грызуны, мозг которых близок к насекомоядным. У грызунов отсутствует четкая дифференциация коры таламуса на специфические и ассоциативные зоны. Известная диффузность представительства сенсорных систем в больших полушариях головного мозга грызунов коррелирует со сравнительно низким уровнем их аналитико-синтетической дея-
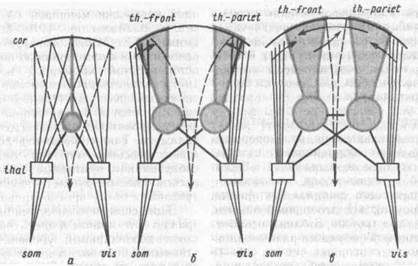
Рис. 70. Схемаэволюционного созревания интегратианыхаппаратов мозга у насекомоядных (а), хищных (б) и приматов (в):
выделены лишь две специфические сенсорные системы — зрительная и соматическая {som и vis); тонкие линии — их проекционные пути; жирные линии и заштрихованные участки—ассоциативные системы, жирные стрелки — кортико-кортикальные связи; пунктирные зоны эфферентных корковых проекций (преимущественно пирамидных); сог кора большого мозга; Thai — зрительный бугор, th. front - таламофронталь-нан; th.-pariet — таламопариетальная ассоциативная система больших полушарий мозга
тельное ти. Слабая выраженность морфологической дифференциации и функциональной специализации полисенсорных структур у грызунов является фактором, определяющим несовершенство интегративной функции мозга.
У белых крыс формирование условного рефлекса на зрительно-слуховой комплекс оказалось также безрезультатным, неоднократные тренировки этой трудной задачи приводили к невротическим срывам. В то же время зрительно-тактильный комплекс выработать удалось, хотя он характеризовался нестабильностью. Примерно аналогичная картина выявилась у кроликов, у которых дифференцирование светового и звукового компонентов от их одновременного предъявления в комплексе не достигало выше 50—60 % уровня. Это отражает низкий уровень аналитико-синтетической деятельности насекомоядных и грызунов.
Второй уровень — хищные, у которых впервые появляются в коре больших полушарий развитые лобные и
теменные ассоциативные поля и соответствующие структуры таламуса. Для хищных характерны существенные структурные и функциональные различия ассоциативных таламокортикаль-ных систем от других мозговых структур. Это наличие сложных нсйронно-синантических компонентов, к которым конвергируют сенсорные потоки, несущие биологически значимую информацию. Причем таламопариетальная система отражает усложнение актов пространственной ориентировки и формирование механизмов, составляющих текущий сенсорный фон для выполнения целенаправленных поведенческих актов. Таламопариетальная система следствие усложнения конструкции и связей зрительной сенсорной системы, она обеспечивает первичной межсенсорный синтез, формирует сложный комплекс обстановочной афферентации и систему «схемы тела».
Таламофронтальная система включается в корковый отдел соматической сенсорной системы с одновременной проекцией на нее лимбических аппара-
тов мозга. Таламофронтальная система участвует в организации предпусковой интеграции и программировании сложных поведенческих актов. Здесь в большей мере отражены процессы мотива-ционно-эмоциональной окраски поведенческих актов благодаря прямым связям этой системы со всем лимби-ческим комплексом. В пределах отряда хищных лобные отделы неокортекса усложняются, увеличиваются их размеры и функциональная роль в организации сложных форм поведения, требующих мобилизации механизмов краткосрочной и долгосрочной памяти.
У хищных (кошки, собаки) выработка условного рефлекса на разномодаль-ный комплекс выполняется за 10—15 опытов. Уровень дифференцирования светового и звукового компонентов от комплекса достигает 70—90 %; но для этого требуется регулярная тренировка. Иными словами, сигнальное значение комплекса поддерживается при условии угашения реакций на отдельные компоненты. Следовательно, у хищных существует более высокий уровень аналити-ко-синтетической деятельности, заключающейся в способности к интеграции разномодальных сигналов в целостный образ.
Другой важной стороной интегра-тивной функции мозга является степень развития процессов памяти и основанное на них свойство прогнозирования предстоящего поведения. В естественных же условиях существования животных любая поведенческая адаптация относительна и имеет вероятностный характер. Поэтому применительно к биологическим условиям обитания животных в вероятностно изменчивой внешней среде имеет место и вероятностное прогнозирование, а значит, адаптивность поведенческих программ определяется степенью их избыточности и подвижности.
Изучалось поведение в стационарных случайных средах. На звуковой сигнал животное должно было направляться к одной из двух кормушек, причем чаще всего к той, где наиболее вероятно получало подкрепление. Степень вероятности подкрепления из каж-
дой кормушки менялась (А. С. Бату-ев, И. В. Малкжова, 1979; А. И. Кара-мян, И. Ь. Малкжова, 1987). Кошки способны к формированию поведения в сторону той кормушки, вероятность пищевого подкрепления из которой наиболее высока. Причем поведение их меняется в соответствии с изменением вероятности подкрепления. Повреждение фронтальных отделов неокор: текса (в основном сенсомоторной коры) сохраняет элементарные условные рефлексы, но разрушает способность к вероятностному прогнозированию.
Процессы межсенсорной интеграции развиты у хищных в достаточной мере, хотя кортикальный уровень ассоциативных систем мозга имеет перекрытие с зонами выхода эфферентных кортикальных трактов. С другой стороны, у хищных достаточно велика роль до-коркового уровня межсенсорной интеграции и кора получает наряду с модально специфичными афферентными залпами уже переработанную неспецифическую импульсацию из ассоциативных ядер. Наконец, каждая из ассоциативных систем у хищных характеризуется доминированием того или иного сенсорного входа, что, естественно, не способствует достижению полной гетеро-сенсорной интеграции.
Третий уровень — приматы, у которых ассоциативные структуры таламуса с их обширной и дифференцированной проекцией в лобные и теменные области коры включаются в состав самостоятельной целостной интегративной системы больших полушарий. Существенной особенностью приматов являются развитые кортикокортикальные связи, с помощью которых ассоциативные поля могут объединяться в целостную иерархически построенную систему. Благода-а ря компактной системе миелинизиро-ванных ассоциативных волокон возрастает роль кортикального уровня взаимодействия специфических сенсорных зон с ассоциативным неокортексом. Ассоциативные поля неокортекса характеризуются тонкой дифференциацией с формированием из нейронных элементов целостных структурно-функциональных ансамблей.
Спецификой ассоциативного неокор-текса является конвергировавшие в нем множества сенсорных сообщений о биологической значимости внешней сигнализации в соответствии с доминирующей мотивацией по независимым друг от друга афферентным каналам. При этом возрастает роль собственно кортикального уровня межсенсорной интеграции и обеспечения процессов краткосрочной памяти. После разрушения либо лобных, либо теменных ассоциативных полей коры не только глубоко страдают процессы межсенсорного синтеза, ориентировочно-исследовательская деятельность, краткосрочная память, но и формирование более простых форм условно-рефлекторной деятельности.
Функциональная значимость отдельных ассоциативных систем расширяется и уточняется в сравнении с хищными. Утрачивается преобладание какого-либо одного сенсорного входа, а следовательно, расширяются возможности их интеграции. Возникает топографическая разнесенность ассоциативных полей от собственно афферентных корковых формаций, что снижает удельное значение сенсомоторной интеграции и расширяет роль коры в осуществлении межсенсорного афферентного синтеза. Возникает все большая их взаимозависимость для обеспечения деятельности целостной интегративной системы полушарий.
Для приматов (низшие обезьяны) выработка условных рефлексов на одновременный комплекс представляет собой относительно легкую задачу, ибо в процессе применения комплексного раздражителя компоненты самопроизвольно утрачивают сигнальное значение и сформированные условно-рефлекторные связи сохраняются месяцами без дополнительной тренировки.'У обезьян можно выработать условный рефлекс даже на трехчленный комплекс из разномодальных сигналов. Это свидетельствует о более высоком уровне аналитико-синтетической деятельности мозга обезьян в сравнении с хищными, что находит отражение и в большей структурной дифференциации и функ-
циональной специализации ассоциативных таламокортикальных систем.
Обезьяны легко справляются с задачами на вероятностное прогнозирование, однако после разрушения в области ассоциативной лобной коры лишаются этой способности, их поведение приобретает однообразный персеверативный характер.
Очевидно, способность использования предыдущего опыта, записанного в долгосрочной памяти для прогнозирования поведения в стационарных случайных средах, претерпевает существенные эволюционные преобразования, которые определяются степенью развития интегративных систем мозга, уровни дифференциации которых коррелируют со степенью совершенства аналитико-синтетической деятельности и организации сложных форм поведения.
Онтогенез ассоциативных систем мозга
Изучение динамики формирования ассоциативных систем мозга показало гетерохронность этого процесса, что, по-видимому, обусловлено включением отдельных звеньев таламокортикальных систем в обеспечение разных поведенческих актов, последовательность созревания которых определяется их необходимостью для осуществления жизненно важных функций новорожденного животного (зрело- и незрело-рождающегося) .
Согласно концепции системогенеза (П. К. Анохин, 1968) неравномерность созревания нервных элементов и связей между ними объясняется их вовлечением в структуру различных функциональных систем. Гетерохронность их созревания определяется значимостью для выживания организма особенно в критические периоды жизни (см. гл. 2), когда новорожденное животное вступает в непосредственное соприкосновение с окружающей средой. Анатомически и функционально первыми созревают те сенсорные механизмы (соматические, акустические), которые обеспечивают выживание на начальных этапах индивидуального развития.
В ассоциативных ядрах таламуса обнаружены проекции в кору больших полушарий уже новорожденных котят (В. П. Бабминдра, Л. А. Васильева, 1987). В то же время неспецифические таламические ядра формируют свои проекции на кору несколько позже, их основная роль состоит в контроле внутриталамической активности.
Несмотря на раннее формирование ассоциативных систем, их окончательное созревание происходит в течение довольно длительного периода и заканчивается у кошек между 2-м и 3-м месяцами жизни, Именно к этому времени у котят впервые формируется полноценный условный рефлекс на одновременный комплекс, приближающийся по своим характеристикам к аналогичному условному рефлексу взрослого животного (Л. А. Васильева, Л. В. Черенкова, 1986).
Когда первый критический этап постнатального развития преодолен, наступает период бурной дифференциации мозговых структур и возникновения между ними двусторрнних связей диффузного характера. Формируется та структурно-функциональная матрица, которая послужит основой для дальнейшего развертывания процессов координации и выделения локальных функциональных структур.
• Таким образом, принцип развития — от диффузного неспецифического к локальному специфическому (А. И. Карамян, 1976) — общебйоло-гическая закономерность, которой подчиняется и динамика развития ассоциативных систем мозга.
Наконец, третий этап связан с моментом формирования тормозных координационных механизмов как в самой коре, так и в глубоких структурах. Появление таких механизмов обеспечивает тонкую специализацию как сенсорных, так и ассоциативных систем мозга, а значит, и различных целостных поведенческих актов.
Дата добавления: 2015-08-04; просмотров: 1788;