Механизмы долговременной памяти
Раньше была распространена точка зрения о том, что механизмы долговременной памяти могут быть сведены к изучению условных рефлексов. Однако детальные исследования показали, что феномен долговременной памяти шире и механизмы, включающие и условно-рефлекторные процессы, разнообразнее и сложнее.
Основанием к этому служит изучение видов памяти животных и человека, проведенное И. С. Бериташвили (1975). Он различал образную, условно-рефлекторную, эмоциональную и словесно-логическую память. Кроме того, у человека психологи и клиницисты выделяют произвольную и непроизволь-
ную память, непосредственное и опосредованное запоминание.
Под образной памятью животных и человека И. С. Бериташвили понимал сохранение в памяти и репродукцию однажды воспринятого жизненно важного объекта. Под эмоциональной памятью понимают воспроизведение пережитого ранее эмоционального состояния при повторном воздействии раздражений, обусловивших первичное возникновение этого состояния (Е. А. Громова, 1980). Эмоциональная память обладает следующими характерными особенностями: 1) она надмодальна, т. с. ее формирование и воспроизведение может происходить при любых сенсорных воздействиях, 2) она формируется очень быстро (в отличие от условно-рефлекторной) и часто с первого раза и 3) она характеризуется непроизвольностью запоминания и воспроизведения информации, т. е. обеспечивает пополнение подсознательной сферы человеческой психики (Э. А. Костан-дов, 1977). Условно-рефлекторная память проявляется в виде воспроизведения условных двигательных и секреторных реакций или заученных привычных движений спустя длительное время после образования. Наконец, словесно-логическая (или семантическая) — это память на словесные сигналы, обозначающие как внешние объекты и события, так и внутренние переживания и свои собственные действия. В любом случае имеет место фиксация предметов и явлений, включающая в свой состав по крайней мере три этапа: I — формирование энграммы, II — сортировка и выделение новой информации и III — долговременное хранение значимой для организма информации.
В отличие от бытовавшем в прошлом представлении о структурных основах долговременной памяти, заключающихся в возникновении новых синапти-ческих контактов между нейронами, канадский ученый Хебб (1949) выдвинул другую гипотезу. Суть ее состоит в том, что фиксация следа памяти связана со стойкими изменениями синаптической проводимости в пределах
определенного нейронного ансамбля. Многочисленные поиски были направлены на выяснение основных факторов, обеспечивающих модуляцию эффективности синаптической передачи и длительное сохранение этих сдвигов в нейронных сетях. В результате этих исследований стало общепризнанным представление о том, что в основе долговременной памяти лежат весьма сложные структурно-химические преобразования как на системном, синап-тическом, так и на клеточном уровнях головного мозга. Ниже будут последовательно рассмотрены основные участники нейрохимических преобразований, обеспечивающие фиксацию памятных следов.
Нейромедиаторные системы.Когда речь заходит о регуляции синаптической эффективности, то естественно внимание обращается прежде всего на системы биологически активных веществ, являющихся посредниками в синаптической передаче. Так, было установлено увеличение содержания свободного ацетилхолина в гипиокампе сразу после обучения оборонительному условному рефлексу. Синтез ацетилхолина, зависящий от вырабатываемых навыков и индивидуальных особенностей животных, стали связывать с процессом консолидации. Весьма чувствительным индикатором холинергической системы мозга является активность ацетилхо-линестеразы (разрушающей ацетилхо-лин), которая закономерно меняется в коре мозга и гиппокампе в зависимости от характера обучения и способности животных к выработке навыков.
Дальнейшие исследования показали, что под влиянием обучения увеличивается количество холинорецепторов, что может быть следствием либо синтеза новых рецепторных молекул, либо демаскировки и активации уже существующих рецепторных белков. Р. И. Кругликов (1986) показал повышение чувствительности кортикальных нейронов, вовлекаемых в формирование условного рефлекса, к микроэлектро-форетически подводимому ацетилхоли-ну. Причем чувствительность возраста-
ет вплоть до фиксации следа в памяти. Э. Кендел (1980) также полагал, что долговременная память связана со стабильным изменением чувствительности к ацетилхолину. Активация рецепторов постсинаптической мембраны облегчает обучение, ускоряет фиксацию, способствует извлечению следа из памяти. И наоборот, антагонисты ацетилхолина нарушают обучение и воспроизведение, вызывая амнезию (Р. Ю. Ильюченок, 1977).
Системы, связанные с образованием и выделением биогенных аминов,— ка-техоламинсргическая и серотонинерги-ческая принимают самое*непосредствен-ное участие в механизмах долговременной памяти (Е. А. Громова, 1980; Р. И. Кругликов, 1981).
Установлено, что обучение животных в моделях с электрокожным подкреплением условных реакций сопровождается активацией норадренерги-ческих систем мозга, а обучение с пищевым подкреплением — снижением метаболизма и уровня норадреналина в мозге животных. Разработана гипотеза, в соответствии с которой нора-дреналин, выделяющийся при подкреплении, пролонгирует активность нейронов, вызванную предъявлением условного стимула, и этим облегчает формирование условного рефлекса.
Значительное снижение норадреналина его антагонистами, разрушение нейронов голубого пятна продолговатого мозга (дающего начало восходящей норадренергической системе) или дор-зального пучка переднего мозга замедляет обучение, вызывает амнезию и нарушает извлечение следа из памяти (Р. И. Кругликов, 1981). Определенную роль играет и дофаминергическая система (Н. Ф. Суворов, В. В. Суворов, 1978), ибо дофамин является предшественником образующегося из него норадреналина.
Значительно более определенная роль в процессах, связанных с консолидацией следов памяти, принадлежит серотонинергической системе мозга. Е. А. Громова (1980) установила, что серотонин ускоряет обучение и удлиняет сохранение навыков, вырабо-
тайных на эмоционально положительном подкреплении, нарушая выполнение и сохранение защитно-оборонительных реакций. Согласно ее концепции моноамины участвуют в процессах обучения и памяти опосредованно, через нейрохимическое обеспечение положительных и отрицательных эмоциональных состояний (см. также гл. 3). Серо-тонин причастен к формированию эмоционально положительных, а норад-реналин — эмоционально отрицательных состояний. Обе моноаминергичес-кие системы находятся в реципрокных отношениях.
В связи с тем, что распространенным приемом исследования памяти является использование различного рода амнезирующих (амнезия — потеря памяти) воздействий стрессорного типа, было выдвинуто представление, что стрессоры действуют на процессы консолидации через нейроэндокринные реакции, оказывая первичное влияние на нсспецифические компоненты этого процесса.
Р. И. Кругликов (1984) разработал гипотезу, согласно которой холинерги-ческие механизмы мозга обеспечивают информационную составляющую процесса обучения. Роль же моноаминер-гических систем мозга сводится к обеспечению подкрепляющих и эмоциональ-но-мотивационных составляющих процесса обучения и памяти. Причем холинергическая система находится под модулирующим влиянием моноаминер-гической системы. Стимуляция норадре-нергических механизмов перестраивает хемореактивные свойства, которые при определенных условиях могут закрепляться, обеспечивая создание и сохранение многонейронной констелляции — энграммы. Если норадренергические механизмы мозга в большей мере при-частны к формированию временных связей, то серотонинергическая — к их фиксации.
X. Матисс (1979) рассматривает взаимодействие обоих типов медиатор-ных систем непосредственно на мембране нейрона (рис. 66). Он полагает, что повторная активация холинерги-ческих синапсов во время стимуляции
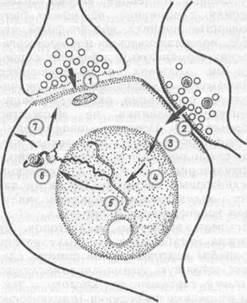
Рис. 66. Механизмы стабильного повышения эффективности синаптической передачи (по X. Ма-тиссу, 1978):
Холинергический медиатор, вызывающий обратимые конформационные изменения субсинаптичеекой мембраны; 2 --- моноамннергический медиатор, активирующий нуклеотндциклазу; 3 циклические нукле-отиды; 4 — активированные протеинкинаэы; 5 -— активация генетического аппарата и изменения синтеза РНК; 6 —- изменение синтеза белков (глюкопротеи-дов); 7 — включение вновь синтезированных белков чолинергическим медиатором изменений субсинап-тической мембраны
вызывает кратковременные конформационные перестройки постсинаптичес-ких мембран, повышающие синапти-ческую проводимость. Если в течение этого времени к нейрону поступают «безусловные» влияния, опосредуемые моноаминергическими системами, то включается цепь внутриклеточных метаболических процессов с участием циклических нуклеотидов. В ходе этих процессов синтезируются полипептиды или белки. Они взаимодействуют с белками постсинаптических мембран, подвергшихся в ходе сенсорной (условной) стимуляции конформационным перестройкам, и стабилизирует эти изменения. В результате нейрон приобретает набор синаптических входов повышенной эффективности, обеспечивающий его участие в составе энграммы.
Данная концепция, весьма правомерная в принципе, не может быть полностью принята, ибо показано, что 95% пресинаптических моноаминерги-ческих терминалей не образуют синапсов на нейронах, а выделяющиеся моноамины, диффундируя на значительные расстояния, оказывают модулирующие влияния на множество образований. Поэтому моноаминерги-ческие системы мозга в последнее время стали причислять скорее к нейро-модуляторным, чем к классическим медиаторным системам. Объектом таких модулирующих влияний является холинергическая система.
Среди вероятных медиаторов, от уровня метаболизма которых зависят процессы долговременной памяти, следует упомянуть гамма-аминомасляную кислоту, глутаминовую кислоту, а также вторичные посредники (циклические нуклеотиды и ионы кальция). Разработка этих вопросов находится только в начальной стадии.
Информационные макромолекулы.Участие нуклеиновых кислот и белков в ключевых процессах обучения и памяти не вызывает сомнений. Теоретические представления на этот счет подразделяются на две группы. Согласно первой группе гипотез обучение и память связаны с кодированием приобретенных форм поведения в информационных макромолекулах. Согласно второй группе гипотез, которая исходит из взаимосвязи гснома и синтеза специфических белков нервной клетки, на основе функционального объединения нейронов возникает структурное их объединение, представляющее собой энграмму памяти.
ф Первая группа гипотез о кодировании индивидуального опыта в макромолекулах базируется на следующих аргументах: качественном изменении РНК и белков при обучении и возможности «переноса памяти» от обученного мозга к необученному с помощью РНК или полипептидов.
X. Хиден (1967) считал, что под влиянием приходящей к нейрону им-пульсации происходит перегруппировка оснований в молекуле РНК, что приво-
дит к синтезу на такой ядерной РНК молекул белка измененной структуры, обусловливающих избирательную чувствительность нейрона именно к данной конфигурации импульсов. Наряду с этим при обучении были описаны синтез полипептидов (Г. Унгар, 1973) и избирательный синтез мозгоспецифи-ческих белков. Много работ, в которых исследуют участие РНК в процессах памяти, не позволяют снять целый ряд принципиальных вопросов и поэтому нельзя исключить неспецифический характер участия нуклеотидов. Так оказалось, что введение животным стимуляторов или ингибиторов синтеза РНК отражается в первую очередь на выработке новых навыков, а не на их сохранении. Пока не получено ни одного убедительного аргумента в пользу признания определяющей роли макромолекул в кодировании индивидуального опыта.
Особо стоит вопрос о так называемом «переносе памяти». Подобного рода исследования на беспозвоночных (планарии, Д. Мак Кеннел, 1959) и млекопитающих (белые крысы, Н. Унгар, 1965) в свое время носили сенсационный характер. Но при их тщательном анализе оказалось, что они содержат ряд методических погрешностей, существенно снижающих их доказательную силу. Вместе с этим нельзя не считаться с достаточно убедительными наблюдениями о наличии стимулирующего влияния экстракта мозга или ликвора обученных доноров на способность к обучению у реципиентов. Видимо, существует какой-то химический фактор, обеспечивающий не прямой «перенос памяти», а облегчающий формирование соответствующего навыка у животных — реципиентов. Г. Адам (1983) на основании своих исследований также приходит к выводу о неспецифическом стимулирующем эффекте экстракта мозга, отвергая за ним функцию «кода памяти».
В последнее время описаны факты прямого переноса условного сахаринового отвращения у крыс (Г. А. Варта-нян, 1986). У одной группы крыс вырабатывали отвращение к сахарино-
вому раствору при сочетаниях питья этого раствора с введением животным хлорида лития, приводящего к интести-нальному шоку. Ликвор обученных животных вводили субокципитально реципиентам, у которых достоверно снизилось потребление сахаринового раствора.
Другой формой «транспорта памяти» является перенос импринтинга у цыплят. Получены некоторые данные, свидетельствующие об олигопеитидной природе вещества-переносчика, а также о быстром включении этого вещества в формирование нового навыка у реципиента (Г. А. Вартанян, М. И. Лохов, 1987).
Для трактовки механизмов «переноса» может быть использована гипотеза об участии иммунологических механизмов в долговременной памяти (И. П. Ашмарин, 1975). Если представить себе, что после прохождения импульсов через синапс усиливается синтез специфических белков-антигенов, то их избыток должен выходить в околоси-наптическое пространство. Эти белки взаимодействуют с рядом расположенными клонами клеток астроцитарной глии и индуцируют их размножение и образование антител. Последние специфически взаимодействуют с постси-наптическими мембранами тех же нейронов и облегчают проводимость в соответствующих синапсах. Данный клон астроцитов сохраняется в течение жизни. В свете данной гиптезы действующим началом «переноса памяти» может быть избыточный антиген пептидной природы, который способен автоматически найти в мозге реципиента либо соответствующую клетку глин, либо синапс.
0 Существует большое число данных, свидетельствующих о том, что полноценный белковый синтез в мозге необходим для процесса консолидации и формирования долговременной памяти. Причем при глубоком угнетении белкового синтеза и относительно кратковременном обучении наблюдается сохранение выработанных рефлексов через минуты или часы обучения. Но уже через часы и сутки после обучения
выявляются глубокие нарушения в сохранении выработанных навыков. Следовательно, процессы белкового синтеза не нужны в ближайшее время после обучения, они понадобятся значительно позже на этапе консолидации энграм-мы. Детальный анализ содержания белков в коре мозга обнаружил, что при формировании новой двигательной координации, связанной с предпочтением одной конечности у крыс, в сенсомотор-ной коре возникает выраженная асимметрия в содержании белков в крупных пирамидных нейронах.
Убедительные данные об участии в функциях памяти получены для двух мозгоспецифических белков S-100 и 14-3-2 (X. Хиден). Первый — активно взаимодействует с мембраной и сократительными белками нейрона при участии нейронов кальция. Второй является ферментативным белком, участвующим в реакциях гликолиза в нейронах. Было обнаружено максимальное содержание в гиппокампе белка S-100 на 4-5-й дни обучения параллельно увеличению ионов кальция. В сравнении с этим Хиден показал, что консолидация памятного следа сопровождается накоплением белка 14-3-2 прежде всего в коре мозга, а не в гиппокампе. Ряд авторов вообще рассматривают S-100 как преимущественно глиальный белок, и лишь белок 14-3-2 связывают со специфическими процессами сохранения эн-граммы. А уровень белка S-100 отражает неспецифическую реакцию мозга на усиленное функционирование церебральных структур.
Для создания устойчивости образованной энграммы должна существовать система обновления специфических рецептор ных белков, котор а я в кл юч ает участки генома, ответственные за синтез соответствующих белков. Либо должны возникать стабильные модификации ДНК, в результате которых в нейроне возникает и поддерживается пожизненно синтез любого нейроспеци-фического белка или, наоборот, выключается необратимо синтез маскирующего белка. Г. Линч и М. Бодри (1984) выдвинули гипотезу, сущность которой состоит в следующем. Повторная им-
пульсация в нейроне сопровождается увеличением концентрации кальция в постсинаптической мембране. Это активирует фермент — кальций зависимую протеиназу, которая расщепляет один из белков мембраны. Его расщепление освобождает замаскированные ранее неактивные белковые глутаматрецепто-ры. Число активных глутаматрецепто-ров возрастает и возникает состояние повышенной проводимости синапса длительностью 3—6 сут.
Эта гипотеза имеет много прямых и косвенных доказательств в свою пользу. Она привлекает внимание тем, что позволяет рассматривать структурную ансамблевую организацию энграммы и при этом учитывает такой важный компонент нейронной конструкции, как дендритные шипики с их белковым цито-скелетом. Аксошипиковые контакты — наиболее пластичное соединение между нейронами, которое может быть ответственно за эффективность синаптичес-кой передачи. Если представить себе, как показали многие авторы, что количество самих шипиков и синапсов на них увеличивается в онтогенезе и прямо зависит от накопления индивидуального опыта, т. е. от образования энграмм памяти, то их использование по поводу новых поведенческих задач и в составе новых обширных констелляций должно подразумевать наличие соответствующих механизмов. С одной стороны, синаптическое соединение с шипиком является структурно достаточно стабильным, ибо сохраняет свою целостность при центрифугировании синапто-сомальных фракций, с другой — функционально весьма подвижным. Последнее может достигаться с помощью механизма изменения диаметра ножки шипика, который, в свою очередь, меняет сопротивление мембраны. Это может обеспечиваться наличием конт-рактильного аппарата в ножке шипика, который представлен молекулами белка актомиозина. Активация этих молекул может возникать при высвобождении ионов кальция из депо, коим является содержащийся в головке шипи-ковый аппарат — эндоплазматичсский ретикулум. Высвобождение ионов каль-
ция происходит при действии медиатора на постсинаптическую мембрану. Сокращение молекулы актомиозина приводит к укорочению и утолщению ножки шипика, в результате чего меняется сопротивление и проведение электрического тока к дендритному стволу. Эти представления носят еще весьма гипотетический характер (А. С. Бату-ев, В. П. Бабминдра, 1984). Значение их в том, что они обращают внимание на построение многонейронной энграммы и на наличие внутрисинапти-ческого цитоскелетного белкового комплекса регуляции синаптической эффективности.
Нейропептиды.Г. Унгар (1977) был первым, кто обратил внимание на участие нейропептидов в явлениях обучения и памяти, которые служат, с его точки зрения, своеобразными маркерами специфических нейронных путей.
В настоящее время основные исследования проведены на гормонах гипо-таламо-гипофизарного происхождения или их фрагментах — коротких цепочках аминокислотных остатков.
Нейропептиды обнаружены в аксон-ных окончаниях нейронов вместе с медиаторами. Установлено, что нейропепти-ды могут усилить или затормозить действие медиатора. Такие нейропепти-ды-спутники вместе с медиаторами создают и поддерживают на постсинаптической мембране специфические ре-цепторные мозаичные наборы, способствующие быстрому проведению определенного вида возбуждения. Пептид-спутник повышает сродство рецептора к основному медиатору, он более стабилен, чем основной медиатор, что обеспечивает пролонгированное облегчение проведения через синапс (И. П. Ашмарин, 1987).
Так, адренокортикотропный гормон (АКТГ) и кортикостероиды и их модификации существенно влияют на обучение и память. Их эффект зависит от интенсивности обучения, он ослабевает по мере увеличения интервала между обучением и введением пептида и является достаточно кратковременным.
Специальные исследования на жи-
вотных с разрушенным гипофизом выявили у них значительный дефицит памяти. Последнее связывали с недостатком в организме гипофизарного гормона — вазопрессина. Дополнительные опыты на линии крыс с генетическим дефицитом вазопрессина и нарушением памяти и улучшением ее после инъекции дополнительных количеств вазопрессина подтвердили участие этого пептида в формировании памятных следов. Причем у этих животных страдал не сам процесс обучения, а именно консолидация сформировавшихся энграмм. Противоположное действие оказывал другой гормон гипофиза — окситоцин, который рассматривают как естественный амнезирующий нейропептид. Он нарушает сохранение выработанных навыков независимо от типа обучения у животных.
Среди других нейропептидов выделяют эндогенные опиоиды — эндорфи-ны и энкефалины, которые оказывают выраженное влияние на обучение и память, замедляют угашение условных рефлексов, улучшают их сохранение, хотя и ухудшают их формирование.
Большинство ученых склоняются к представлению о том, что нейропептиды регулируют память через взаимодействие с медиаторами и через их влияние на метаболизм макромолекул. Действительно, оказалось, что предварительное разрушение голубого пятна предотвращает усиливающее действие вазопрессина на процессы консолидации. Такой же эффект возникает при предварительном разрушении серото-нинергических нейронов ядер шва. Но в отсутствие норадренергической системы чувствительность серотонинергичес-кой системы возрастает. В свою очередь имеются данные о том, что влияние серотонина на консолидацию энграммы опосредуется через систему опиоидных пептидов.
Таким образом, прослеживается тесная взаимосвязь всех без исключения нейрохимических механизмов обучения и памяти. По-видимому, изменение белкового метаболизма является тем ко-
нечным звеном, через которое реализуются любые воздействия на процессы обучения и памяти. За счет изменений белкового метаболизма осуществляются процессы формирования и закрепления многонейронной энграммы. «Хранителем всех форм нейрологичес-кой памяти служит система межнейронных взаимодействий, и участие информационных биополимеров проявляется лишь во включении или выключении активности различных участков предшествующего генома, а не в синтезе новых нуклеотидных или аминокислотных последовательностей»1. Дальнейшая задача состоит в разделении специфических и неспецифических факторов формирования энграммы памяти с углубленным анализом ее нейрохимических и структурных основ.
Различают генетическую, иммуноло-гическую и нейрологическую (нервную) формы памяти. Последняя подразделяется на кратко- и долговременную память, помимо которых выделяют еще и промежуточную память. Перевод значимой информации из кратко- в долговременную память называется консолидацией энграммы, т. е. образованием структурно-химических изменений, фиксирующих внешнюю ситуацию и отношение к ней самого субъекта.
Кратковременная память только что минувших событий или впечатлений, извлеченных из долговременной памяти, основывается на импульсной реверберации в замкнутых нейронных цепях. Долговременная память формируется на основе синтеза макромолекул — нуклеиновых кислот и белков — и связана с активацией генетического аппарата нервной клетки, в результате чего возникают изменения в мембранах нейронов и межнейронных связях.
1 Ашмарин И. П. Молекулярные механизмы нейрологической памяти//В кн.: Механизмы памяти (Руководство по физиологии). Л., 1987. С, 61.
Глава
Дата добавления: 2015-08-04; просмотров: 2685;