МЕХАНИЗМЫ ПАМЯТИ
Можносчитать, что организация сложных форм поведения — это прежде нечто организация работы механизмов памяти.
(М-Р. Гаазе-Рапопорт и Д. А. Поспелов)
Виды и формы памяти
Формы биологической памяти.Многие из специалистов придерживаются представления о памяти как сложном феномене фиксации, сохранения и воспроизведения информации о взаимодействии между объектами. Это определение включает широкий диапазон феноменов от элементарной памяти неживой природы до психонервной памяти человека. Принципиальное отличие живого от неживой природы состоит в структурном воспроизведении живого, что в принципе невозможно, если живая система не запомнит свою стабильную организацию. Следовательно,
Биологическую память можно определить кик способность живых существ (или их популяций), воспринимая воздействия извне, закреплять, сохранять н в последующем воспроизводить вызываемые этими воздействиями изменения функционального состояния н структуры
(И. П. Ашмарин 1975). Стабилизация органических систем и их структурное самовоспроизведение опирается на генетическую память, как память биологического вида. Носителем генетической памяти являются нуклеиновые кислоты, которые позволяют обеспечить стабильность хранения информации. В основе изменчивости такой информации в филогенезе лежит мутагенез. Воспроизведение измененных форм происходит с участием белков-ферментов. Ге-
нетическая детерминация свойств высшей нервной деятельности рассмотрена в гл. 3.
Второй формой биологической памяти, развивающейся позже в ходе эволюции, является иммунологическая память, которая тесно связана с генетической памятью. В самой общей форме иммунологическая память состоит в способности после первой встречи с генетически чужеродными телами и веществами узнавать их при повторной встрече, связывать и включать неспе-цифическис механизмы их уничтожения. Такие чужеродные вещества именуются антигенами, а иммунные белки, обладающие способностью разрушать антигены, называются антителами. Основными участниками иммунного ответа являются иммуно компетентные лимфоциты. Их поверхностная мембрана оснащена определенным набором антител. Причем взрослый организм снабжен набором генетической информации для синтеза всего разнообразия антител. Эти антитела на лимфоцитах служат рецепторами для антигенов. Каждый лимфоцит имеет рецепторы к одному или к нескольким сходным антигенам. А все лимфоциты, несущие одинаковый рецептор, принадлежат к одному клону, т. е. являются потомками одной материнской клетки с таким же рецептором.
Процесс селекции клонов и обусловливает иммунологическую память. Первая встреча с антигеном вызывает увеличение числа соответствующих лимфоцитов (формирование клона) и их диффсренцировку на эффекторные клетки и клетки памяти. Если первые живут несколько дней, то вторые остаются в организме на всю жизнь и при повторной встрече с антигеном способны вновь превращаться в клетки обоих типов. Селективными агентами, которые обеспечивают материал для отбора, являются антигены. Они «узнают» рецепторы антител, связываются с ними и стимулируют их размножение. Значит, из огромного набора вариаций антител антиген отбирает единственную и стимулирует ее количественный рост. Эта форма памяти представляет собой эволюционное развитие генетической памяти в сторону ее большей гибкости и индивидуальной реактивности (Г. А. Вартанян, М. И. Лохов, 1987).
Одним из основных свойств нервной системы является длительное хранение информации* о событиях внешнего мира и реакциях организма на эти события, а также использование этой информации для построения текущего поведения. Эти свойства и легли в основу формирования нервной, или нейрологи-ческой памяти. В эволюции она возникла в связи с дифференциацией нервной системы и оказалась самой сложной по проявлениям и механизмам.
В каждый момент времени животные и человек переходят из своего прошлого к новому, ранее не испытанному состоянию, и их поведение в настоящем в значительной мере определяется совокупностью накопленного опыта в прошлом. Вот эту непрестанно возрастающую совокупность следов от пройденного, определяющую поведение в наступающем настоящем, и называют памятью (А. А. Ухтомский). Таким образом, память как результат обучения заключается в таких изменениях в нервной системе, которые сохраняются в течение некоторого времени и существенно влияют на характер протекания будущих рефлекторных реакций. Ком-
плекс таких структурно-функциональных изменений, включающий запечатле-ние не только определенной внешней ситуации, но субъективное отношение организма в ней, получил название процесса образования энграммы. Используя весь ранее приобретенный опыт, а следовательно, опираясь на факторы, уже отсутствующие в настоящем, энграмма оказывается избыточной. Благодаря своей избыточности энграмма служит основой активности организма и реалистического прогнозирования им будущих ситуаций.
Общепризиапо представление о системной организации памяти, т. е. ее организации во времени и пространстве. И изучение элементарных механизмов памяти всегда должно ориентироваться на целостное понимание этой функции мозга. Одним из трудных вопросов является отражение в памяти фактора времени. Известно, что в памяти события размещаются по оси времени с отражением их реальной длительности. Ясно, что формирование и воспроизведение таких энграмм возможно лишь при допущении временного «свертывания» событий на основе существования собственного времени мозга. На основе такой компрессии времени создается внутренний кронотоп (по А. А. Ухтомскому), т. с. внутренний пространственно-временной образ внешнего единого пространственно-временного мира. При воспроизведении такой эпграммы неизбежно отражение реальной, физической встречи с объектом. 11оэтому энграмма из модели прошлого перемещается в будущее и становится фактором, определяющим цель предстоящего поведенческого акта. Значит, цель в виде реальной энграммы предшествует следствию, т. е. действию с его результатами (Р. И. Кругликов, 1987).
Временная организация памяти.После электрического ответа рецеитор-ной клетки на внешнее воздействие возникают следовые процессы, продолжающиеся некоторое время уже при отсутствии реального раздражителя. Эти первичные следовые процессы составляют основу сенсорной памяти.

Рис. 57. Виды и формы памяти (но Г. М. Чайченко, 1987)
Длительность хранения следов в сенсорной памяти не превышает 500 мс, стирание следа осуществляется за 150 мс. Так, зрительный образ сохраняется во время мигания, при чтении, восприятии речи и пр. На этом же виде памяти основано слитное восприятие изображений в кино и телевидении. Предполагают, что сенсорная память человека не зависит от его воли и не может быть подвергнута сознательному контролю. Длительность хранения следов в сенсорной памяти продолжает оставаться дискуссионной. В частности, требуют объяснения способности некоторых людей-эйдетиков, у которых период сохранения зрительного образа может достигать десятки минут.
Одна из классификаций видов памяти приведена на рис. 57. Следующий за сенсорной памятью период, связанный с хранением информации, именуется краткосрочной памятью. И наконец, наиболее значимая информация хранится в долгосрочной памяти. Такое деление видов памяти принимается большинством исследователей, хотя некоторые и предлагают свою более дробную классификацию. Кратко-и долговременная память могут различаться по своим механизмам и оказывать друг на друга взаимное влияние. Консолидация энграммы, т. е. ее переход из кратковременной в долговременную, — принципиально важный мо-
мент физиологии и биохимии памяти. Некоторые полагают, что периоду консолидации соответствует особый вид промежуточной памяти, которая как бы встраивается между долговременной и кратковременной памятью.
Существует мнение о том, что кратковременная и долговременная память представляют собой последовательные этапы единого процесса. В то же время ряд гипотез исходит из точки зрения о параллельном развитии процессов кратко- и долговременной памяти. Предполагают, что кратковременная (нестабильная) память образуется за счет конформанионных перестроек макромолекул и реализуется с участием си-наптического и сипаптосомального уровней регуляции (рис. 58). Перемещение ионов и/или кратковременные метаболические сдвиги во время синап-тической активности могут привести к изменению эффективности синаптичес-кой передачи, длящейся миллисекунды и секунды. Большинство ученых усматривают в основе кратковременной памяти прежде всего электрофизиологические механизмы, связанные с многократным циркулированием импульса-ции (реверберацией) по замкнутой системе нейронов. С помощью психологических тестов Эббингауза было установлено, что объем кратковременной памяти человека измеряется 7±2 единицы, т. е. бессмысленные слова
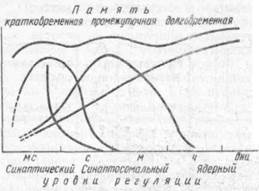
Рис. 58. Временная организация., памяти(по X.Матиссу, 1978)
после однократного их предъявления воспроизводятся испытуемым лишь в количестве 7±2.
Синаптосомальный уровень включает конформационные изменения структурных и ферментных белков, изменения концентрации и перемещение нейромедиаторов. Это и есть длящаяся минуты и часы промежуточная память. Промежуточная память, как полагают психологи, расширяет объем кратковременной памяти и увеличивает ее длительность. Долговременная память формируется на основе синтеза макромолекул — нуклеиновых кислот и белков.— и связана с активацией генетического аппарата клетки. Следовательно, в отличие от предшествующих процессуальных видов памяти долговременная память представляет из себя новую внутри мозговую функциональную структуру, базирующуюся в результате на изменениях в мембранах нейронов и на межнейронных связях.
Благодаря, в основном, исследованиям психологов в пределах краткосрочной памяти выделена первичная память, связанная с мысленным повторением материала с целью запоминания и его интерпретации (рис. 57). Длительность этой памяти ограничивается несколькими секундами, и материал стирается при его замене на новый. В свою очередь, долговременную память подразделяют на вторичную путем создания ассоциации, т. е. взаимоотношений между отдельными элементами и явлениями, которые могут хра-
ниться минуты и годы, и третичную память, куда входят навыки, постоянно сопровождающие жизнь человека: способность к чтению, письму, профессиональные навыки и пр., которые практически не забываются. Кроме того, различают у человека произвольное и непроизвольное запоминание; образную, моторную, логическую, эмоциональную, условно-рефлекторную памяти. Конечно, нельзя не видеть, насколько еще далеки друг от друга физиологические и психологические схемы классификации видов памяти. Накопленные к настоящему времени физиологические и биохимические данные позволяют с определенной достоверностью выделить три вида памяти: а) сенсорную, б) кратковременную и в) долговременную. .
§ 22. Механизмы кратковременной • памяти
Нейрофизиологические корреляты краткосрочной памяти.К данной категории или стадии относится память на только что минувшие события. Представим себе следующую экспериментальную ситуацию. Обезьяна обучается по условному сигналу — зажигание лампы справа или слева на пульте — выполнять движение нажатия на соответственно правую или левую педаль или рычаг. Если выбор педали или рычага выполнен правильно, такое действие вознаграждается пищей {А. С.Ба-туев и др., 1988). Но сигналы подаются в случайном порядке и после каждого сигнала наступает пауза, которая может длиться разное время — от 5 до 20 с. В течение этого периода, называемого периодом отсрочки, животное не имеет доступа к рычагам и педалям: они скрыты от него непрозрачным экраном. После отсрочки экран открывается и обезьяна может выполнить требуемое действие (рис. 59). Значит, животное, получив условный сигнал, должно удержать в памяти информацию о его местоположении в течение всего периода отсрочки, а затем использовать эту информацию для осуществления правильного движения.
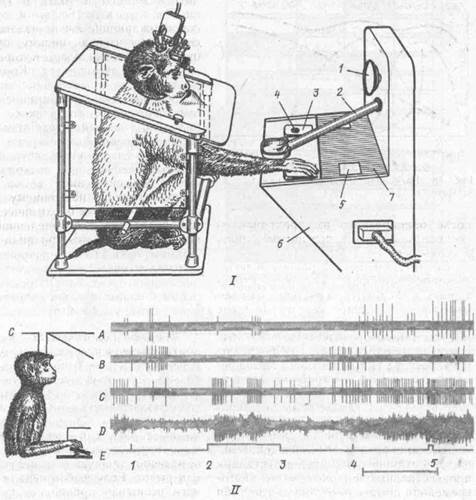
Рис. 59. Методика изучения нейронной активности мозга в тестах на краткосрочную память у обезьян;
/ — экспериментальная процедура; // -- экспериментальная программа и ее нейрональные корреляты; / — лампа-вспышка (предупреждающий сигнал); 2 — тубус для подачи пищи; 3 — педаль; 4 — лампа (условный сигнал); 5 — прозрачное окно в экране (7); 6 — общая панель; А и В активность нейронов лобной коры; С — двигательной коры; О — электромиограмма дельтовидной мышцы плеча, Е — отметки сигналов (/ — предупреждающий; 2 и Л — условный; 4 — пусковой (отодвигание экрана); 5 — нажатие на клавишу). Калибровка времени - 2 с
Такое отсроченное поведение является общепризнанным методическим подходом для изучения механизмов кратковременной памяти, с помощью которой сохраняется информация о только что прошедших событиях, но необходимых для выполнения предстоящего поведенческого акта. У таких животных из различных отделов мозга с помощью микроэлсктродоп длительно
регистрируется импульсная активность нескольких нейронов. Исследования показали, что информация о пространственном расположении условного сигнала кодируется в импульсной активности нейронов прежде всего лобной и теменной ассоциативных полей коры больших полушарий. Такое кодирование осуществляется либо рисунком разряда нейронов, либо частотой им-

Рис. 60. Сравнительное'распределение пространственно селективных нейронов лобной и теменной коры обезьян в тестах на кратковременную память:
цифры показывают процентное отношение к общему числу пространственно селективных нейронов. Этапы экспериментальной программы: / — предупреждающий сигнал; 2 — период неспецифического ожидания; 3— условный сигнал; 4 — отсрочка; 5 — пусковой период программы (открытие акра на, нажатие на рычаг, получение пищевого подкрепления)
пульсации нейронов, получивших название пространственно селективных. Оказалось, что эти нейроны не представляют собой однородную популяцию, а подразделяются на несколько групп (рис. 60). Часть нейронов обнаруживает свойство пространственной селекции только в период предъявления условного сигнала — такие нейроны названы сенсорными. Другая группа нейронов обнаружила различия в импульсной активности только п период отсрочки — их именуют нейронами памяти. Третья группа аналогичное свойство проявила только после открытия экрана, в момент, непосредственно предшествующий моторному акту, поэтому их назвали нейронами моторных программ. Описаны и смешанные типы нейронов.
Детальный анализ самого периода отсрочки показал, что все ее время заполняется кратковременными вспышками импульсной активности разных
нейронов лобной коры: в начале отсрочки, в середине и в конце, либо вспышек активности одних и тех же нейронов. И только незначительное число клеток поддерживает высокую импульсную активность на протяжении всего периода отсрочки (рис. 61). Пространственно селективные сенсорные нейроны стоят на входе сложных' нейронных кортикальных цепей и лишь кодируют сенсорные признаки сигнала, в данном случае его пространственное положение. Нейроны, активирующиеся в начальный период отсрочки, детектируют полученную информацию о свойствах условного сигнала, далее следует передача возбуждения по принципу эстафеты другим нейронным популяциям» каждая из которых отдает аксоны к нейронам-сумматорам, и от последних происходит передача задержанной таким образом информации на программирующие и пусковые структуры.
Именно для лобных отделов коры, где и обнаружены вышеописанные феномены, характерны замкнутые нейронные круги типа «ловушек», в которых импульсный поток может циркулировать (реверберировать) до тех пор, пока не будет переключен на эфферентные модули из крупных пирамид (рис. 62). Последнее может служить структурной основой удержания п ловушках импульсных потоков и эстафетного переключения активации от одного нейронного объединения к другому. Лоренте де Но был одним из первых морфологов, кто описал сложные замкнутые цепи нейронов в разных частях мозга. На основе морфологи ческих данных была создана модель памяти из замкнутых цепей, в которых импульсы циркулируют без подкрепления (рис. 63). И. С. Беритов (1948) показал, что часть звездчатых клеток проекционных зон коры имеет аксон, заканчивающийся в поле своих дендри-тов. Такая структура может работать как ловушка возбуждения.
Внутрикорковую реверберацию рассматривают как базовый механизм краткосрочной памяти для лобной коры. Важнейшим фактором формирования
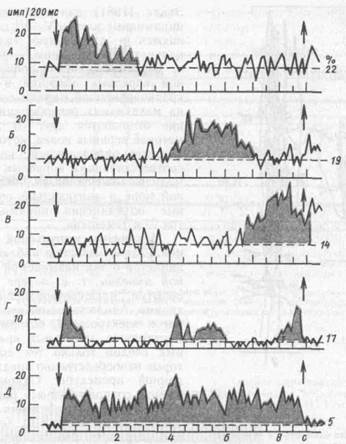
Рис. 61. Основные типы перестроек импульсной активности нейронов лобной коры в период отсрочки (по А. А. Пирогову)
Дата добавления: 2015-08-04; просмотров: 2329;