Конвергентная теория формирования временных связей
Разработке основных законов центральной нервной координации физиология обязана исследованиям Ч. Шер-рингтона, сформулировавшего фундаментальный закон в работе нервной системы —«принцип общего пути». Если афферентный нейрон является индивидуальным или частным путем, то эфферентный нейрон является общим путем для импульсов, возникающих в любом из многочисленных источников рецепции. Подобная координация складывается на основе морфологического соотношения афферентных и эфферентных путей, где первые всегда значительно преобладают над вторыми. Так формируется и реализуется механизм конвергенции — схождения множества афферентных стимулов в единственный анатомически ограниченный эфферентный канал. Частные пути, конвергируя между собой, соединяются в промежуточные пути и окончательно конвергенция завершается общим конечным путем в виде воронки. Каждый этап, на котором два или более афферентных нейрона конвергируют на третьем нейроне, который по отношению к ним является эфферентным, предполагает возможность влияния или столкновения их влияний.
Идею о принципе конвергенции творчески развил А. А. Ухтомский (1954), распространив его на всю центральную нервную систему, на все ее этажи. Он полагал, что в результате конвергенции наступает интерференция стимулов, ко-
торая реализуется в определенный нервный акт. Отсюда становится понятным богатое развитие синапсов и концевых ветвлений с тенденцией увеличить поверхность соприкосновения нервной клетки с окончаниями приходящих аксонов.
Автор принципа доминанты выдвинул новые положения об интегрирующей деятельности нервной системы. Принцип статической конвергенции путей по отношению к исполнительному пути при дальнейшей разработке расширяется в принцип подвижной и физиологической конвергенции нервных импульсов по отношению к области преобладающего текущего действия (принцип доминантны).
А. А. Ухтомский находил тесную связь между учением Шеррингтона об общих путях и теорией И. П. Павлова об условных рефлексах. В статье «Об условно-отраженном действии» он писал: «Если воронка Шеррингтона имела в виду постоянные рефлекторные дуги, закрепившие функциональную и морфологическую связи между собой наследственно и филогенетически, то И. П. Павлов улавливал самое закладывание и новообразование связей в этой воронке, привлечение все новых рецептивных сфер к конвергенции относительно эфферентных (исполнительных) приборов»1.
Конвергенция гетеросенсорных стимулов была положена в основу понимания механизмов образования условных рефлексов. Так, Дж. Экклс (1953)
^Ухтомский Л. А. Доминанта. Л., 1966. С. 175.
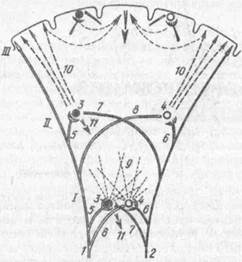
Рис. 41. Схима образованияусловного рефлекса (по А. Гасто,. 1958):
/ — ретикулярная формация нижней части ствола мозга; // — таламическая ретикулярная формация; /// --кора мозга; / — афферентный путь безусловного раздражителя; 2 — афферентный путь условного раздражителя; 3 и 4 — ретикулярные нейроны; 5 и 6 — коллатеральные волокна афферентных путей, вызывающие возбуждение ретикулярных нейронов; 7 и 8 - коллатеральные волокна афферентных путей, не вызывающие возбуждения нейронов вследствие непроходимости синапсов; 9 и 10 — восходящие пути от ретикулярной формации, активирующие кору диф-фузно (9) и локально (10), И безусловно-рефлекторная реакция. Временная связь образуется между афферентными влияниями условного и безусловного раздражителей
предположил существование нслокали-зованного «центра конвергенции», где сходятся условный и безусловный раздражители. Встреча их в центре конвергенции, взаимодействие и интеграция завершаются формированием особой пространственно-временной структуры разрядов, которая затем передается в нейронную сеть. Многократные сочетания включают механизмы потенциации путей условного сигнала, который приобретает способность запускать условно-рефлекторный акт. Длительность хранения этого эффекта обеспечивается механизмами реверберации возбуждения в нервных сетях. А. Фессар (1962) показал, что конвергенция, т. е. сближение в пространстве мозга разномодальных сигнализаций, является важнейшим условием формирования временной связи на
уровне отдельных нейронов. В качестве двух других условий он выдвинул взаимодействие этих сигнализаций на нейронах и использование следовых процессов. Взаимодействие между конвергирующими сигналами связано с длительным удерживанием следов активности. В этих процессах важную роль играют механизмы тонической и посттетаниче-ской потенциации.
Ф Таким образом, конвергенция и потенциация образуют некую общую цепь динамических процессов, связывающих интеграцию в пространстве (обусловленную конвергенцией) с интеграцией во времени (обусловленную постепенным нарастанием облегчения на окончаниях аксонов) (А. Фессар, 1964).
Первичная конвергенция осуществляется преимущественно на клетках ретикулярных образований, таламуса и ствола мозга и передается на корковые нейроны в интегративной форме («проецируемая мультивалентность»). Корковые структуры регулируют конвергентные свойства ретикулярных структур, что обеспечивает специализацию условного рефлекса (рис. 41).
Упомянутые концепции не могли объяснить свойственные условному рефлексу устойчивость и приспособи-тельную пластичность, ибо не придали должного значения фактору биологического подкрепления. Именно это было учтено П. К. Анохиным (1968), который считал, что решающим фактором в механизмах образования условных рефлексов выступает биологическое, значение подкрепляющего рефлекса. Конвергирующие на нейронах сенсорные и биологические модальности определяют организацию условного рефлекса — его сигнальные свойства и приспособительное значение. Стимулы различной сенсорной и биологической модальности воздействуют при сочетаниях на синаптические входы нейронов с гетерохимической организацией этих входов. Свойства подкрепляющего фактора определяются специфическими химическими реакциями, которые он вызывает, и соответствующими молекулярными преобразованиями в преде-
лах мембраны и цитоплазмы нейрона. Здесь и происходит «сцепление» условной и безусловной стимуляции с последующим выходом сложившейся интеграции на аксон в виде клеточного разряда.
Само понятие «сцепление» следов условного и безусловного стимулов предполагает наличие специфических ультраструктурных и молекулярных преобразований в цитоплазме нейрона. Гипотетическая схема, предложенная М. Я. Рабиновичем (1975), приведена на рис. 42. Сочетание стимулов двух сенсорных модальностей (6\ и б^) приводит к образованию кратковременной связи типа сенсо-сенсорной ассоциации. Сочетание же каждого из сенсорных стимулов с биологически значимым подкреплением (а) эмоциональ-но-мотивационного характера приводит к выработке устойчивой связи, обладающей свойствами условного рефлекса.
Признавая, что наиболее существенные процессы образования временных связей реализуются на молекулярном уровне нейрона, следует иметь в виду, что системные механизмы временных связей всецело опосредованы сложными нейрофизиологическими преобразованиями.
Выделяют три типа проявлений мультисенсорной конвергенции (Л. Л. Воронин, 1974). Первым типом является «специфический ассоциативный», при котором клетки ствола мозга, тала-муса или коры реагируют специфическим ответом на стимулы разных сенсорных модальностей. Вторым типом является «специфический проекционный», при котором клетка реагирует специфической реакцией на адекватный для данной системы раздражитель и неспецифической — на стимулы других модальностей. Наконец, третий тип именуется неспецифическим и характеризуется неспецифической ответной реакцией нейронов на разномодальные стимулы. М. Я- Рабинович (1975) разработал гипотетическую схему, в которой синоптические входы для условного и безусловного стимулов конвергируют на нресинаптической («вход-
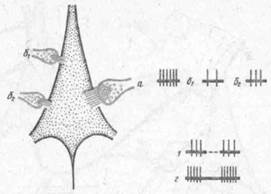
Рис. 42. Вероятная схема взаимодействия синап гических входов с гетерохимической организацией для стимулов биологической (а) и сенсорных б1] и б2) модальностей (по М. Я Рабиновичу,
1975):
различные свойства входов для сенсорных и биологически значимых сигналов условно изображены в виде размеров синапса и его активных зон
ной») клетке в цепи нейронов. Сложившаяся ассоциация («интеграция») передается на постсинаптический нейрон («выходной»), но по одному общему пути и для сигнального и для подкрепляющего раздражителей (рис. 43).
При рассмотрении разномодальной конвергенции необходимо учитывать чрезвычайно важное свойство нейронов и нервных сетей, а именно способность к организации пластического характера конвергенции. В этой связи следует разграничивать конвергенцию анатомическую, или статическую, и конвергенцию функциональную, или динамическую. Особенности организации нервных сетей головного мозга обеспечивают связь по принципу «от каждого ко всем». Следовательно, морфологические предпосылки могут создать такие условия, когда на одном нейроне будут конвергировать самые разнообразные влияния. В процессе эволюции и в динамике постнатального онтогенеза в головном мозге закрепляется олигосинаптическая сеть, обеспечивающая четкий адресат проведения информации наряду с системой полиси-наптических сетей, связи между компонентами которой могут носить функциональный временный характер. Это об-
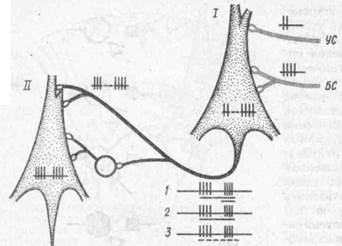
Рис. 43. Гипотетическая схем передачи условной связи, с.т жившейся в процессе сочетаний с «входного нейрона» (/) на «вы ходной» нейрон (//) по одного общему пути (по М Я Рубин'.. вичу, 1975):
клетка круглой фирмы — вставочный нейрон возбуждающего действия; на осциллограммах — различ ные формы условной 'активности. которые возможно регистрировать с «выходного» нейрона: / — обычная реакция на условный н безусланный стимулы (УС и БС), обозначен ные длинной н короткой линией пол осцилограммой; '2 — воспроизведение следоных разрядов на БС при его пропуске в сочетании; 3 воспроизведение следовых разрядов на УС и БС при пропуске всего сочетании (обозначено пунктиром) в условиях реакции на «время*
стоятельство, т. е. высокая вариативность свойств полисенсорных нейронов, и обусловливает многообразие их реакций на разномодальную стимуляцию.
Дата добавления: 2015-08-04; просмотров: 2455;