Клеточные аналоги условного рефлекса
Выделяют две основные гипотезы обучения — синоптическую, предполагающую, что механизмом формирования условных рефлексов является изменение эффективности синапсов, и мембранную, утверждающую, что основным механизмом условного рефлекса является изменение свойств возбудимой постсинаптической мембраны. Как мы увидим далее, такое деление весьма условно, однако представляется удобным для первого знакомства с проблемой.
Нервные механизмы обучения обеспечиваются благодаря двум основным свойствам нервной системы — реактивности и пластичности. Если реактивность — это способность отвечать на раздражения, то пластичность — это способность изменять реактивность под влиянием последовательных раздражений. В спою очередь реактивность определяется возбудимостью постсинаптической мембраны и эффективностью соответствующих синапсов. Возбудимость нейрона измеряется силой тока, необходимой для генерации по-
тенциала действия, а синаптическая эффективность — амплитудой моноси-наптического постсинаптического потенциала.
Уже давно высказывалось предположение . о том, что в основе обучения лежит изменение эффективности межклеточных контактов или проторе-ние новых контактов. На рис. 44 приведены основные схемы. Простейшим является синапс Дж. Экклса (А), повышающий свою эффективность после собственной активации (фасилита-ция). Такие синапсы могут служить для объяснения феноменов посттетаниче-ской потенциации. Дополнением к ним являются синапсы с постепенным снижением эффективности при повторной активации (Б). Они могут обеспечивать эффекты габитуации и дегабиту-ации. Как мы уже отмечали, при выработке условных рефлексов необходимо временное совпадение пресинап-тической активации с активацией подкрепляющих систем, которые могут оказывать также пресинаптическое действие (В). Д. О. Хебб (1959) главным условием синаптической модификации считал совпадение пресинаптической активности с разрядом постсинаптиче-ского нейрона (Г). Затем к этому была добавлена модель Альбуса, связанная не с увеличением эффективности синапса, а с ее снижением (Д). Наконец, в качестве синтетической схемы
(Е) рассматриваются случаи изменений синаптической эффективности с участием трех факторов -- сенсорного притока, активации подкрепляющей системы и достижения результата.
Вопрос о характере преобразований нейрональной активности в динамике выработки условного рефлекса начал исследоваться в связи с внедрением в практику физиологического эксперимента тонких методов микроэлектродного анализа и последующей компьютерной обработки импульсных реакций нейронов. Попытки использовать внутриклеточную регистрацию постси-нантических электрических потенциалов привели к необходимости проводить исследования либо на относительно простых нервных сетях, либо на центральных нейронах высших млекопитающих с максимальным упрощением требуемых для анализа межнсйронных отношений. Такие упрощенные модели принято рассматривать в качестве клеточных аналогов условного рефлекса.
Группой венгерских физиологов (О. Фехер, А. Барани, 1981), разработана модель ассоциативного обучения, при котором сигнал меняет свое действие под влиянием последующего безусловного эффекта.
Рассмотрим процедуру обусловливания при «условном» раздражении вент-ролатералыюго ядра таламуса (VL) и антидромной стимуляции пирамидного тракта («безусловный стимул»). Сначала, как показано на рис. 45, электрическая стимуляция VL с частотой 1/с сопровождается только подпороговыми ПСП. Последующее антидромное раздражение пирамидного тракта вызывает один потенциал действия. По мере сочетаний раздражения таламуса с антидромной стимуляцией амплитуда и/или длительность ТПСП увеличиваются и остаются таковыми в течение нескольких минут. Амплитуда ВПСП изменяется также, но к 50-му сочетанию ВПСП достигает порога генерации и «условный» сигнал начинает вызывать 1 —2 потенциала действия. Эффект антидромного раздражения при этом не меняется. К 100-му сочетанию амплитуда ВПСП еще боль-
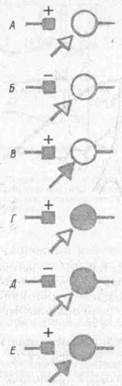
Рис. 44. Основные виды гипотетических модифицирующихся синапсов {по Л. Л. Воронину, 1986):'
А — активации синапса, Б — ю же; В — активации синапса и подкрепляющей системы; Г — активация синапса и постсинаптнческого нейрона; // — то же; £ активация синапса; постсинаптического нейрона и подкрепляющей системы; квадрат — модифицирующиеся синапсы, стрелка — синапсы подкрепляющей системы, кружок — постсинаптический нейрон, зачерненные знаки элементы, активация которых входит в условие модификации данною синапса, ( + ) — повышение, ( — ) — снижение эффективности синапса
ше возрастает, и число потенциалов действия достигает трех. Если проводить угашение, т. с. подавать одно «условное» раздражение, не сопровождая его антидромной стимуляцией, то амплитуда ВПСП и число генерируемых потенциалов действия уменьшается, и спустя 350 изолированных раздражений таламического ядра ответная «условная» реакция исчезает. Внутриклеточная поляризация исследуемого нейрона, приводившая к генерации потенциалов действия, усиливала эффект раздражения вентролате-
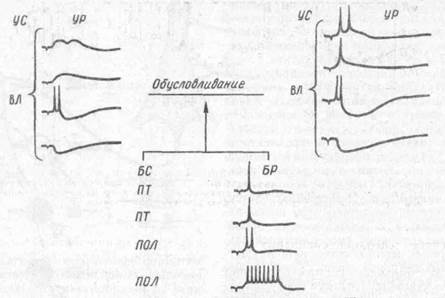
Риг.45. Эффекты гетеросинаптического облегчения на корковом нейронепирамид ного тракта у кошек (по А Барани и О Фехеру. 1980):
УС — условный стимул; УР — условная реакция; БС — безусловный стимул; БР — безусловная реакция; слева — до сочетания; справа — после сочетания. Приведены постсинап-тические реакции нейрона на адекватную (ВЛ), антидромную (ПТ) пирамидного тракта стимуляцию и внутриклеточную поляризацию (ПОЛ) постоянным током
рального таламуса. Однако ни многократное применение только «условного» стимула, ни псевдообусловливание (процедура применения условного и безусловного раздражителей в случайном порядке) не привели к генерации потенциалов действия исследуемого нейрона. Уровень мембранного потенциала и сопротивление мембраны при выработке временной связи менялись по-разному в различных клетках. Это позволило думать об определяющей роли при выработке временной связи избирательных синаптинеских сдвигов. Было показано, что положительные и дифференцировочные связи с исследуемым нейроном опосредуются разными синапсами независимо друг от друга, Е. Н. Соколов (1981) справедливо полагает, что элементарным ло-кусом пластического изменения является не нейрон, а синапс. Речь идет о синапсе Хебба, который селективно изменяется, если за синаптической посылкой следует спайковый разряд нейрона. Такое явление получило наимено-
вание гетеросинаптического облегчения.
Этот же феномен использовался для объяснения ряда эффектов обусловливания в другой модели (Л. Л. Воронин, 1982). Синаптические аналоги условного рефлекса создавались в зоне коркового представительства передней конечности бодрствующего кролика сочетаниями электрического раздражения двух точек коры. Условным стимулом служило подпороговое (неэффективное) раздражение коры на некотором расстоянии от исследуемого коркового нейрона, а безусловным — пороговое для постсинаптической реакции раздражения поверхности коры над исследуемым корковым нейроном. При активирующих формах условной связи в ответ на условный стимул возникали В11СП, амплитуда которых постепенно возрастала до критического уровня генерации потенциалов действия (рис. 46). При отмене подкрепления, т. е. при у га теп ни, амплитуда ВПСП постепенно уменьшалась с пре-
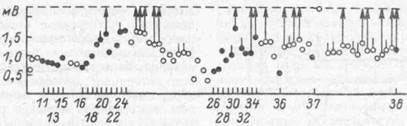
Рис. 46. Усиление ВПСП и импульсных реакций на аналоге корково-корковогоусловного рефлекса (по Л. Л. Воронину, 1986);
По оси ординат— изменение ам'плитуды ВПСП в ходе опыта; по оси абсцисс — номера сочетаний; светлые кружки — амплитуда ВПСП в ответ на изолированный УС; темные кружки то же, во время сочетаний; вертикальные черточки н стрелки отмечают генерацию частичного и полномерного потенциалов действия соответственно
крашением генерации спаиковых разрядов.
При тормозных формах временной связи условный стимул постепенно вызывал развивающееся угнетение механизма генерации импульсных разрядов, которое не сопровождалось существенной гиперполяризацией мембраны клетки или достоверным уменьшением ВПСП. При этом установлено, что дифференцирование одним и тем же нейроном подкрепляемых и неподкрепляемых стимулов реализуется различными синаптическими входами и поддерживается благодаря механизмам гетеросинаптического облегчения.
Одной из моделей аналога временной связи на микропопуляции нейронов коры больших полушарий является микроионофоретическая аппликация медиаторов (глутамата и ацетилхоли-на), используемых в качестве аналогов ассоциируемых раздражителей (Б. И. Котляр, 1986). Установлено, что отдельные нейроны микропопуляции в радиусе действия медиаторов обнаруживают специфические ассоциативные перестройки своей активности. Последние могут заключаться в следующем: изменения реактивности к сигнальному раздражителю, воспроизведение реакций на подкрепляющий раздражитель в ответ на действие условного сигнала и реакция на время в границах действия ассоциируемых раздражителей.
В. С. Русинов (1969) пришел к вы-
воду, что суммационный рефлекс и доминанта представляют собой начальные стадии образования любого условного рефлекса и сами по себе могут служить удобной моделью для изучения простых форм временной связи. Им была разработана модель корковой поляризационной доминанты, состоящая в том, что поляризация постоянным током участков сенсомоторной коры кролика при действии ранее неэффективных раздражителей — световых вспышек, приводила теперь к ритмическому сокращению передней конечности (корковая проекция которой поляризовалась) в такт световой стимуляции. Установившаяся под влиянием поляризации новая форма реагирования на раздражитель сохраняется до 30 мин после выключения тока.
Эта модель послужила существенным толчком для выявления пластических характеристик корковых нейронов. Установлено, что поляризация слабым постоянным током увеличивает активность полисенсорных нейронов, что является одним из непременных механизмов возникновения движения на ранее индифферентный раздражитель. Причем функциональные и морфологические сдвиги происходят не только в пирамидных нейронах, но и в интернейронах и глиальных клетках, которые также подвергаются поляризации.
Все основные свойства доминанты: повышенная возбудимость, стойкость возбуждения, способность к суммиро-
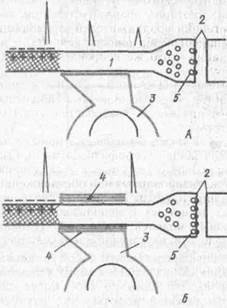
-Рис. 47. Схема процесса образования времен ной связи (по Л. И. Ройтбаку, 1972):
А —- «потенциальный» синапс; К — образование временной связи (превращение «потенциального» синапса в «актуальный»); / — пресинаптический участок нервного окончания; 2 — синашическая щель; 3 --- олигодендроцнт; 4 — ммелиновая оболочка; 5 — медиатор в синаптических пузырьках. Эффективность проведении тока по терминальному волокну увеличивается вследствие формирования вокруг него оболочки из олигодендроиита
ванию возбуждений, инерции, торможению и растормаживанию — были обнаружены на вышеописанной модели. В доминантном очаге происходит установка на определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данных условий ритм работы, когда этот очаг становится наиболее отзывчивым и тормозятся другие работающие очаги. Иными словами, образуется единый функционально организованный ансамбль или система. Подчеркивается особая роль нейронов нижних слоев коры, которые дольше сохраняют ритм раздражения, чем клетки верхних слоев. Обращается серьезное внимание на существование в системе базальных дендритов пирамидных клеток дендро-дендритических контактов, которые рассматриваются как важ-
нейший субстрат межнейронных отношений. В. С. Русинов полагает, что наиболее адекватным отражением функционального состояния системы нейронов является уровень их постоян ного потенциала, а следовательно, электротонические влияния имеют наиболее существенное значение для образования временной связи. Поляризационный очаг, созданный постоянным током, служит удачной моделью для изучения поляризационно-электротони-ческой гипотезы формирования доминанты и простых форм временных связей (В. С. Русинов, 1969).
Специальное внимание к нейрогли-альным взаимоотношениям при формировании клеточных аналогов условного рефлекса привлек А. И. Ройтбак (1969). Он выдвинул глиальную гипотезу, согласно которой в основе образования и укрепления временных связей лежит миелинизация центральных аксонов, возбуждающихся от условного раздражения. При действии безусловного раздражения в соответствующих областях коры происходит деполяризация глиальных клеток: возбуждение большого числа нервных элементов импульсами из специфических и неспецифических путей приводит к выделению r межклеточные щели и аккумуляции ионов калия. Автор полагает, что деполяризация олигодсндроциток служит сигналом к образованию миелина; условием миелинизации является совпадение во времени деполяризации олигодендроцитов и физико-химических изменений в прилегающих немиелини-зированных пресинаптических термина-лях после их предшествующего возбуждения (рис. 47).
В последние годы широко развернулись исследования механизмов временных связей на животных с относительно простой нервной системой. Наиболее удобными для экспериментирования оказались моллюски. Простейшей моделью служил полностью изолированный нейрон висцерального ганглия виноградной улитки. Локальная аппликация ацетилхолина сочеталась с внутриклеточной инъекцией деполяризующего тока, в результате было по-
казано, что изолированные нейроны моллюсков (т. е. лишенные своих сетевых свойств) не способны формировать ассоциативный ответ в виде воспроизведения безусловной реакции сигнальным раздражителем.
Клеточные механизмы образования условного рефлекса на морском моллюске аплизии изучались Э. Кенделом (1980). Исследовался оборонительный рефлекс: условный сигнал — умеренной силы раздражение сифона, вызывающее втягивание жабры в мантийную полость, безусловный раздражитель — сильная стимуляция хвоста. В результате сочетаний защитная реакция на стимуляцию сифона усиливалась. Удобство этой модели состояло в том, что был точно определен нейронный состав рефлекторной дуги, в которую входят 24 сенсорных нейрона, образующие прямые связи с интернейронами и 6 моторными нейронами, иннервирующими жабру и сифон. Стимуляция хвоста возбуждает нейроны, контактирующие с иресинаптическими окончаниями сенсорных нейронов.
В данной модели обнаружились явления сенситизации, привыкания и иейрогуморального (субклеточно-молекулярного) воздействия безусловного раздражителя на всех этапах выработки временной связи Суммация заключалась в неспецифическом гетероси-наптическом облегчающем эффекте — увеличении ВПСП, связанном с увеличением выброса медиатора из пресинап-тического окончания. Суммация этих процессов приводит к возникновению длительной деполяризации, составляющей сущность проторения путей при образовании временной связи. Процессы привыкания связаны с ограничением потоков кальция в пресинапти-ческие окончания, что приводит к гиперполяризации нейронов и снижению их ответа на безусловный раздражитель. Нейрогуморальный эффект безусловного раздражителя достигался благодаря механизмам регуляторных нейропептидов (см. гл. 6).
Э. Кендел (1980) считает, что в основе условной реакции втягивания жабры у аплизии лежит механизм пре-
синаптического облегчения (сенситизации), который формируется при конвергенции раздражителей, приводящих к спайковой активности нейрона, и активации этого же нейрона через модулирующий вход от безусловного раздражителя. Аналогичный механизм допускается и у позвоночных животных, хотя прямые доказательства этого отсутствуют.
ф Какими бы информативными для задач общей нейрофизиологии ни были аналитические подходы к исследованию наркотизированных или обездвиженных препаратов позвоночных и беспозвоночных животных с использованием различных моделей и аналогов условных рефлексов, их конечной целью является расшифровка тех феноменов мульти-нейронной активности, которые регистрируются на бодрствующем мозге при выполнении животным того или иного целостного условно-рефлекторного акта. Сегодня мы еще не можем использовать весь арсенал сведений аналитической нейрофизиологии для понимания клеточной активности бодрствующего мозга. Различия двух основных подходов базируются на степени проявления свойства пластичности, а именно — высокой изменчивости реакций одного и того же нейрона у бодрствующего животного и значительно большей стабильности реагирования нервных клеток у наркотизированных препаратов.
Дата добавления: 2015-08-04; просмотров: 1668;