Доминанта определяет вероятность возникновении той или иной рефлекторной реакции в ответ на текущее раздражение.
Как общий принцип работы нервных центров доминанта подчиняется определенным закономерностям. Прежде всего в нервных сетях живого организма почти непрерывно возникают потоки возбуждений из разнообразных центральных и периферических источников. Это физиологически оправдано, ибо в организме наблюдается значительное преобладание рецепторов, улавливающих тончайшие изменения внешней и внутренней среды, над соответствующими центрами в головном и спинном мозге и над исполнительными органами.
ф А. А. Ухтомский сформулировал следующие основные свойства доминанты: 1) повышенную возбудимость; 2) стойкость возбуждения; 3) инертность возбуждения; 4) способность к суммированию возбуждения. К этому следует добавить способность доминанты переходить в тормозное состояние и вновь растормаживаться (В. С. Русинов, 1969). Ну и, конечно, доминанта только потому и может стать
таковой, что одновременно с созданием и подкреплением самое себя она приводит к сопряженному торможению центров антагонистических рефлексов.
Что касается повышенной возбудимости, то отнюдь не сила возбуждения в центре, а именно способность к дальнейшему увеличению возбуждения под влиянием нового приходящего импульса может сделать центр доминирующим. Далее А. А. Ухтомский подчеркивает важное свойство доминирующего центра достаточно интенсивно, продолжительно и стойко накоплять и поддерживать в себе возбуждение, что приобретает значение господствующего фактора в работе прочих центров. Все это создаст предпосылки для хронического свойства доминанты, ее инертности. Наконец, относительно суммации возбуждений А. А. Ухтомский высказался весьма категорично, полагая, что судьба доминанты решается тем, будет ли центр способен суммировать свои возбуждения под влиянием доходящих до него импульсов или они застанут его неспособным к суммированию.
Определяющую роль при формировании доминантного очага играет состояние развивающегося в нем стационарного возбуждения, в основе которого лежат различные физико-химические процессы. Определенный уровень стационарного возбуждения чрезвычайно важен для дальнейшего хода реакции. Если этот уровень возбуждения мал, то диффузная волна может поднять его до состояния, характерного для доминанты, т. е. создать в нем повышенную возбудимость. Если же уровень возбуждения в центре и без того высок, то при приходе новой волны возбуждения возникает эффект торможения. По доминантный очаг не следует рассматривать как «центр сильного возбуждения». Определяющую роль в этом процессе играет не количественный, а качественный фактор — повышенная возбудимость, т. е. отзывчивость на приходящие волны возбуждения и способность центра суммировать эти возбуждения. Именно такой центр — наиболее возбудимый, отзывчивый и впечатлительный в дан-
ный момент -- отвечает на приходящий стимул, анатомически даже не относящийся к нему, именно такой центр, первым вступая в работу, предопределяет на какой-то более .или менее продолжительный промежуток времени новый ход реакции. Считая одной из основных черт доминанты ее направленность (векторность), А. А. Ухтомский вместе с тем отмечал, что векторная направленность произвольного поведения является результатом доминанты.
Не менее важную роль в процессе формирования доминанты играет процесс сопряженного торможения. Состояние возбуждения в центре, подкрепленное возбуждениями из самых отдаленных источников, будучи достаточно стойким, т. е. инертным, в свою очередь снижает способность других центров реагировать на импульсы, имеющие к ним прямое отношение. В случае доминант низшего порядка сопряженное торможение рассматривается как следствие возникновения определенного фокуса возбуждения, тогда как в случае доминант высшего порядка (например, кортикальных) они являются прямым фактором объединения «очагов» возбуждений в некоторый рабочий ансамбль. Но торможение в других центрах наступает лишь тогда, когда возбуждение в формирующем нервном центре достигает достаточной величины. Именно сопряженное торможение играет наиболее важную роль при формировании доминанты, и это торможение должно быть своевременным, т. е. иметь координирующее значение для работы других органов и организма в целом. Координационное торможение — это процесс энергетически более емкий, «дорогой», позднее вырабатывающийся, чем простое возбуждение. Торможение — это наиболее тонкий, интимный процесс и вместе с тем наиболее ранимый.
Сложные рефлекторные акты осуществляются не одним анатомическим центром, а несколькими центрами, расположенными в различных отделах спинного и головного мозга и образующими своеобразную функциональную
констелляцию центров в работе целостного мозга. Стационарное возбуждение в констелляции центров представляется в виде определенного цикла, когда каждый участник этой констелляции, будучи связан с соседними и стимулируя их, в свою очередь получает от них стимуляцию. Таким образом складывается своеобразная «циклическая машина», которая долгое время сохраняет в себе возможность вспыхивать и усиливаться под влиянием текущих раздражений. Состояние возбуждения в доминанте А. А Ухтомский назвал ее симптомокомп-лексом.
Для образования доминантной установки необходимо нарастание в элементах констелляции состояния возбуждения еще до того, как возникнут дискретные волны возбуждения, способные к проведению и суммированию своих эффектов, т. е. когда слагаемые будущей констелляции заработают воедино, когда сложится их рабочий ансамбль.
Доминантный процесс — не привилегия определенных этажей нервной системы: он может складываться в любой центральной группе в зависимости от условий подготовки, развития возбуждения в ней и сопряженного торможения антагонистических механизмов. Процесс торможения не только оформляет и поддерживает доминирующую констелляцию, но может привести и к прекращению ее функционирования, т. е. сама доминанта может тормозиться.
Часто сравнивая физиологический механизм с механизмом техническим, А. А. Ухтомский отмечал не только то, что он образуется по ходу самой реакции, но и то, что в отличие от технического механизма он имеет не одну, а множество степеней свободы, дающих ему возможность осуществлять и множество переменных операций. В самой природе субъекта наличие конвергенции обусловливает наделение существующей системы исполнительных органов множеством степеней свободы, которые обеспечивают новые функциональные разновидности, достигающие-
ся новыми сочетаниями прежних ана-томофизиологических данных.
Выбор одной степени свободы при сопряженном торможении других достигается посредством доминанты, направленной на определенный рабочий вектор,
При рассмотрении вопроса о превращении доминанты из симптомокомплек-са в общий механизм действия, поведения и жизнедеятельности организма А. А. Ухтомский нередко использовал понятие «функционального центра», «функционального органа». Этим подчеркивалось следующее: дело не столько в том, что в нервной системе складываются определенные констеля-ции связанных между собой центров, морфологически далеко разбросанных по мозговой массе, а в том, что они функционально объединены единством действий, своей векторной направленностью на определенный результат. В понятие органа может быть включено всякое сочетание сил, могущее привести при прочих равных условиях всякий раз к одинаковым результатам. Орган — это прежде всего механизм с определенным однозначным действием.
Громадное разнообразие морфологических черт того или иного образования нашего тела впервые приобретает значение физиологического органа после того, как открывается значение этих морфологических деталей для рабочих слагающих однозначно определенной физиологической равнодействующей
(А. А. Ухтомский, 1950).
Единство действия достигается способностью данной совокупности приборов взаимно влиять друг на друга с точки зрения усвоения ритма, т. е. синхронизации активности нервных центров. В этих процессах огромную роль играет усваивание темпа и ритма возбуждений в центрах, входящих в состав констелляции. Каждый из потенциальных компонентов констелляции изначально обладает своим определенным темпом и ритмом возбуждений. При формировании же единой констелляции, этого созвездия созвучно работающих ганглиозных участков, взаимно совозбуждающих друг друга, определяющую роль играет увязка во времени, скоростях, в ритмах действия, т. е. в сроках выполнения отдельных моментов реакции. Именно в констелляции
более всего возможно возникновение ритмического процесса. Происходит как бы своеобразное развертывание констелляции, когда постепенное «сыгрыва-ние» ритмов в отдельных ее компонентах приводит к достижению некоторого общего «марта» возбуждения в ней. При этом сопряженное торможение прочих центров, не входящих в состав данной констелляции, будет развиваться тем более, чем полнее констелляция будет участвовать в ритмической работе. Процесс синхронизации темпов и ритмов активности нервных центров, входящих в состав определенной констелляции, делает ее доминирующей.
ф Ее доминирование, следовательно будет зависеть от того, насколько полно она будет вовлечена в дело, насколько наладившийся ритм возбуждений будет осуществляться без перебоев, подкрепляясь новыми текущими раздражениями, насколько при этом будут сопряженно тормозиться другие рефлексы.
Основным аппаратом управления ритмами в тканях и органах, по А. А. Ухтомскому, является кора головного мозга, обеспечивающая обоюдный процесс подчинения темпов и сроков жизнедеятельности темпам и срокам сигналов из внешней среды, равно как подчинение сроков в постепенно освояемой среде потребностям организма. Одновременно с поддержанием синхронизации ритмов в рабочей констелляции кора мозга принимает участие в вовлечении и других нервных центров в этот процесс синхронизации.
ф Одной из черт доминирующей констелляции является способность ее отдельных компонентов или центров принимать участие в других рефлекторных актах, входя в состав соответствующих им констелляций. Но как и чем определяется подобный переход? С одной стороны, сохранение того или иного компонента — участника рабочей констелляции — зависит от того, насколько стойко и продолжительно он способен сохранять в себе темп и ритм, навязанный ему данной констелляцией, т. е. от его инерции. С другой стороны, переход данного нервного центра в
работу другой констеляции будет зависеть от того, насколько быстро данный центр способен усвоить темп и ритм нового межцентрального рабочего цикла, возникшего в нервной системе.
Постепенно формируясь, рабочая констелляция проходит несколько стадий. Сначала доминанта в центрах вызывается ее непосредственным раздражителем под влиянием внутренней секреции, рефлекторными влияниями и пр. На этой стадии, названной стадией укрепления наличной доминанты, по преимуществу, она привлекает к себе в качестве поводов к возбуждению самые разнообразные внешние раздражения. В констелляцию посредством генерали-зованного возбуждения наряду с необходимыми для данного акта нервными центрами вовлекаются и посторонние клеточные группы. В этом достаточно неэкономичном процессе проявляется диффузная отзывчивость доминирующей констелляции к различным раздражениям. Но постепенно, в процессе повторного осуществления данного рефлекторного акта, диффузная отзывчивость сменяется избирательным реагированием только на те раздражения, которые создали данную доминанту. Подбор биологически интересных рецепций для доминанты (стадия выработки адекватного раздражителя для данной доминанты и вместе с тем стадия предметного выделения данного комплекса раздражителей из среды) приведет к образованию новых адекватных поводов для той же доминанты. Теперь выполнение доминантного акта происходит более экономично, ненужные для него нервные группы затормаживаются. При повторении доминанты репродуцируется и свойственный только ей единый ритм возбуждения.
Восстановление доминанты по кортикальному следу может быть эскизным, т. е. более экономичным ее воспроизведением через кору. При этом комплекс органов, участвующих в переживании восстановленной доминанты, может быть сокращенным и ограничиться только кортикальным уровнем. Полное или эскизное возобновление доминанты возможно лишь в том слу-
чае, если возобновится хотя бы частично раздражитель, ставший для нес адекватным. Это обусловлено тем, что между доминантой как внутренним состоянием и данным комплексом раздражителей была установлена прочная адекватная связь. На этой стадии доминанта возобновит подбор новых, «биологически интересных поводов для себя». Поэтому при кортикальном воспроизведении доминанты, которое является очень подвижным сочетанием ее кортикальных компонентов, происходит интеркортикальное обогащение новыми рецепциями кортикальных же компонентов доминанты.
Что же представляет собой кортикальный след, по которому доминанта может быть пережита заново? Это своего рода интегральный образ, своеобразный продукт пережитой ранее доминанты, в котором в единое целое сплетены соматические и эмотивные признаки доминанты с ее рецептивным содержанием, т. е. с комплексом раздражений, с которым она была связана в прошлом. При создании интегрального образа важную роль играют как периферические, так и кортикальные компоненты. Интегральный образ — это своеобразная памятка пережитой доминанты и вместе с тем ключ к ее воспроизведению с той или иной степенью полноты.
Если доминанты восстанавливаются по своим кортикальным компонентам, т. е. более экономично, как «мимолетное воспоминание», причем с малой инерцией, то при новых условиях они всегда обходятся с помощью прежнего опыта. Биологический смысл эскизно протекающих доминант состоит в том, чтобы «... по поводу новых и новых данных среды очень быстро перебрать свой арсенал прежних опытов для того, чтобы из них путем очень быстрых их сопоставлений избрать более или менее идущую к делу доминанту, чтобы применить ее к новому заданию. Целесообразность или нецелесообразность выбранной доминанты прошлого решает дело»1. Если же доминанта
1 Ухтомский А. А. Доминанта. Л., 1966. С. 258.
восстанавливается почти с прежней полнотой, которая предусматривает оживление работы всей соматической констелляции, то она переживается с большей инерцией, занимая более или менее продолжительный период жизне^ деятельности организма. Подбирая при этом вновь биологически интересные для себя раздражения из новой среды, доминанта по новым данным переиите-грирует старый опыт.
Корковый очаг стационарного возбуждения.А. А. Ухтомский экспериментально электрическим или фармакологическим путем формировал в спинном и головном, мозге очаг стационарного возбуждения, в результате чего, он обнаружил, что одним из его свойств является способность реагировать на посторонние для него возбуждения.
В лаборатории В. С. Русинова (1967) было показано, что стационарное возбуждение в коре головного мозга сопровождается сдвигом уровня постоянного потенциала. Эти же сдвиги могут быть вызваны искусственно путем поляризации коры слабым постоянным током. Анодизация слабым постоянным током коркового представительства одной из конечностей бодрствующего кролика создает условия, при которых ранее индифферентные световые и звуковые раздражения начинают вызывать двигательную реакцию соответствующей конечности. Специальными опытами было показано, что такой очаг возбуждения обладает не только способностью суммировать возбуждение, но и свойствами интерности, торможения при чрезмерных возбуждениях и восстановления при оптимальных условиях раздражения.
Структурами, принимающими активное участие в образовании такого очага, являются апикальные дендриты пирамидных клеток, интернейроны и клетки глии. Их анодизация создает условия, при которых значительная часть нейронов коры становится полисенсорной.
ф Таким образом, в доминантном очаге происходит установка на определенный уровень стационарного возбуждения, способствующий суммирова-
нию ранее подпороговых возбуждений и переводу на оптимальный для данных условий ритм работы, когда этот очаг становится наиболее отзывчивым и тормозятся другие работающие очаги.
Однако события, происходящие в поляризационном очаге, характеризуют лишь одну из сторон процесса складывания доминанты. Оказалось, что в формировании коркового доминантного очага участвуют различные неспецифические образования таламуса. Дополнительная поляризация мезэнцефали-ческих и таламических ядер ретикулярной формации облегчала формирование двигательной доминанты, усиливала ее. Если корковый доминантный очаг тестируется световой стимуляцией, то дополнительная оптимальная поляризация наружного коленчатого тела подкрепляет доминанту.
На «поляризационной модели» исследовались механизмы суммации путем сочетания подпороговых раздражений сенсомоторной коры кошки и световых вспышек. Замечено появление в результате суммации движения соответствующей конечности (Р. А. Павлы-гина, 1982). Этому предшествует изменение конфигурации дендритного потенциала той же зоны коры — появляется дополнительная волна, связанная с тормозным процессом. Сразу вслед за этим наблюдается «отдача» в виде учащения нейронной активности, совпадающей с волнами послеразряда и движением конечности животного. Следовательно, одним из механизмов суммации является двухфазная реакция с Непосредственным переходом от торможения к возбуждению. Эти эффекты суммации исчезают сразу после прекращения сочетаний. Однако, если те же сочетания проводить на фоне поляризации коры слабым постоянным током, они сохраняются в виде следа десятки минут после прекращения раздражений.
Сама же поляризация приводит к изменению как фоновой активности, так и формы реагирования нейронов: учащение реакции на световое раздражение появилось у ранее нсреа-гировавших нейронов, увеличилась ин-
тенсивность ответа у ранее реактивных клеток коры, тормозная реакция на раздражитель сменилась на возбудительную. Была показана возможность двух функциональных состояний доминантного центра. Первое длительное, скрытое, без выхода на эффектор, с повышением возбудимости не только в поляризационном участке сенсомоторной коры, но и в других районах, способствующее иррадиации возбуждения из любого участка к доминантному очагу. Второе — в момент двигательной реакции угнетаются ответные реакции коры на дистантные раздражения, что является проявлением сопряженного торможения при реализации двигательной доминанты.
Повышение возбудимости участка коры под влиянием поляризации не только обусловлено изменениями мембранного потенциала эфферентных пирамидных нейронов, но и само влияет на другие структуры. К таковым, в частности, относятся вставочные клетки и глия: увеличивается количество олигодендроцитов около пирамид и интернейронов, возникают сдвиги содержания РНК в цитоплазме и ядрах клеток. Кроме того, меняются и нейронные взаимоотношения, которые рассматриваются как морфологический коррелят их высокой функциональной активности.
Описывая свойства доминанты как системы центров с высокой возбудимостью, А. А. Ухтомский подчеркивал значение временного фактора ее существования. Именно на длительности функциональных сдвигов в центрах и базируются такие свойства доминанты, как стойкость, инертность, способность к суммации. Иными словами, доминанта опирается в своем существовании на наличие определенного набора оптимальных раздражителей, но в основном на следовые процессы, которые и отличают доминанту от частных коротких рефлексов. В противовес такому консервативному началу доминанты, которое выражается в подкреплении ее за счет всевозможных поводов и впечатлений, А. А. Ухтомский подчеркивал ее прогрессивную роль в процессе новообра-
зования рефлексов на среду, когда происходит быстрый подбор «нужных, имеющих непосредственную связь впечатлений».
Гипоталамический очаг стационарного возбуждения.С целью анализа естественных событий в организме рассмотрим свойства отдельных хеморецеп-тивных зон мозга. Такие «центральные хеморецепторы» или их совокупности описаны, обнаружена их специализация к определенному химическому соединению или гормону. Специфические хеморецепторные нейроны, высокореактивные к колебаниям «физиологических констант» организма, описаны в большом числе в ядрах гипоталамуса. Поэтому первичный очаг возбуждения при формировании некоторых естественных биологических доминант (голода, жажды) может возникать именно в этих образованиях мозга. Инертность такого очага обеспечивается за счет длительной активации соответствующих низкопороговых хеморецепторных нейронов и окружающих их глиальных клеток.
Например, стойкая мотивация жажды сопровождается возникновением в супраоптическом и в латеральном ядрах гипоталамуса длительных вспышек гиперсинхронизованной активности, которые с усилением мотивации иррадии-руют на кору больших полушарий (рис. 67). Эти вспышки провоцируются также световыми и звуковыми стимулами и исчезают после внутриполостного введения воды или внутривенного введения физиологического раствора. Уже эти признаки свидетельствуют о том, что при мотивации жажды в гипоталамусе формируется стойкий доминантный очаг (А. С. Батуев, Б. Г. Гафуров, 1988).
Подобного рода первичные очаги стационарного возбуждения можно рассматривать в качестве одного из систе-мообразующих факторов при формировании доминирующей нейродинами-ческой констелляции мозговых структур.
Анализ нейронной активности латеральных ядер гипоталамуса при формировании мотивации жажды (А. С. Батуев, Ю. Ли, 1987) обнаружил возник-
новение центральных и периферических перестроек во вкусовой сенсорной системе. Мотивация жажды повышает возбудимость входных рецепторных элементов вкусовой сенсорной системы и облегчает проведение к центральным нейронам информации о химических качествах пищи путем раскрытия рецептивных полей нейронов и перехода их на режим широкополосного приема.
Возникновение таких первичных очагов в свою очередь может вести к появлению вторичных и более мощных очагов в других отделах мозга, где имеются предпосылки для продолжительного удержания следовых процессов. Например, в нейронах гиппокампа посттетаническая потенцияция длится часами или даже сутками.
Другой структурой, где могут формироваться вторичные очаги стационарного возбуждения, является кора головного мозга. Принципиальным отличием таких очагов от первичных является исходная причина их возникновения. Так, если первичный очаг в гипоталамусе возникает и поддерживается в первую очередь за счет гуморальных факторов, то вторичные очаги в коре больших полушарий являются проекционными по своему происхождению и поддерживаются за счет широкой конвергенции к ним множества суммирующих здесь сенсорных влияний. По-видимому, следовые процессы здесь настолько мощны, что приводят к каким-то структурным изменениям, длительно модифицирующим передачу в синапсах. Одним из возможных механизмов являются стационарные изменения цито-скелета в аксошипиковых синапсах.
Если применительно к биологическим мотивациям локализацию первичных очагов можно связывать с гипо-таламическими ядрами, то топография вторичных очагов должна опираться на карту гипоталамо-кортикальных связей. Р. А. Павлыгина (1982) считает, что в нормально функционирующем организме должны быть заложены механизмы, допускающие сосуществование нескольких доминант одновременно. Сопряженные тормозные взаимовлияния характерны для функцио-
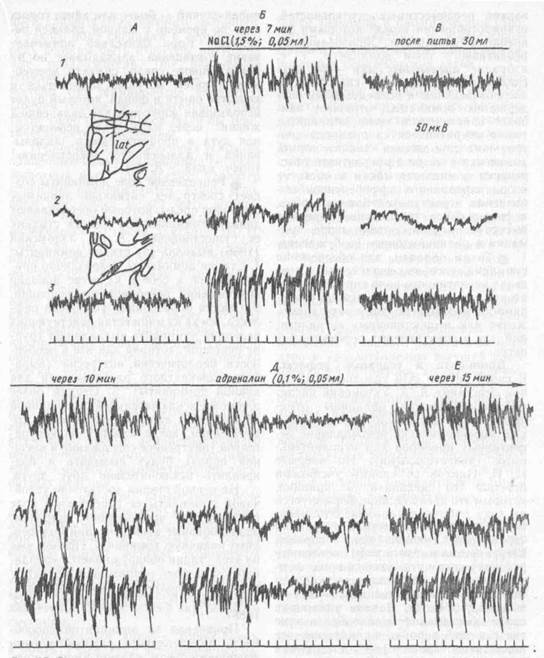
Рис. 67. Суммарная биоэлектрическая активность коры и ядер гипоталамуса у кошек с мотивацией жажды, созданной прямым введением хлорида натрия в латеральные ядра гипоталамуса (по 5. Г. Гафурову и И. В. Якунину, 1989):
А — исходный фон; Б — после однократного введения хлорида натрия (горизонтальная линия сверху) появляется высоковольтная активность и поисковое поведение в сторону поилки; В ~- после питья 30 мл воды оба признака жажды исчезают; Г — спустя 10 мин жажда восстанавливается, Д — иновь исчезает после введения в латеральный гипоталамус адреналина; Е — через 15 мнн поведенческие и биоэлектрические признаки жажды восстанавливаются; / — латеральный отдел гипоталамуса; 2 — сенсомоторная область коры; 3 — супраоптичес-кое ядро гипоталамуса; вертикальная стрелка - введение адреналина. Калибровка: время — 1 с, амплитуда 50 мкВ.
нально несовместимых , еятельностей, пзаимоотношсния мсжд ' которыми закреплены генетически. Образование же рецептивного поля доминанты, т. е. избрание адекватные для себя сиг-гнальных раздражений, способных к воспроизведению и последующему поддержанию доминанты, отражает наиболее пластическое звено доминанты, также опирающееся на процессы центрального торможения. Значит, торможение как в части афферентного обеспечения доминанты, так и в части ее координированного эфферентного выполнения играет определяющую рол*, и сопряженное торможение первично выступает в роли организатора сум-мации в наличном очаге возбуждения.
Ф Таким образом, для обеспечения суммации сенсорных воздействий в пределах рецептивного поля кортикального вторичного «очага» повышенной возбудимости необходимо допустить торможение как второстепенных сигнализаций, так и конкурентных рефлекторных актов.
Доминанта и условный рефлекс.В своей работе «Об условно-отраженном действии» А. А. Ухтомский писал: «Шсррингтоновские принципы отмечают особо как успело отпечатлеться уже морфологически преобладание рецептивных приборов над исполнительными (двигательными). Но впервые И. П. Павлов со своими учениками осветил тот механизм и принцип, которым это преобладание формируется на ходу работы организма. Это механизм «условного рефлекса» и принцип «временной связи». Если воронка Шеррингтонаимела в виду постоянные рефлекторные дуги, закрепившие функциональную и морфологическую связи между собой наследственно и филогенетически, то И. П. Павловулавливал самое закладывание и новообразование связи в этой воронке, привлечение все новых рецептивных сфер конвергенции относительно эфферентных (исполнительных) приборов..- Связь исполнительного органа с новыми рецептивными поводами может закладываться, так сказать, случайно, вследствие того, что работа данного исполнительного органа
имела случай — более или менее совпадать во времени с данной дальней рецепцией. Кора бдительно примечает такие совпадения, закладывает по их поводу новые связи, сначала временные, а затем могущие закрепиться в качестве опыта и фонда, который будет использован животным для дальнейшей жизни... перед нами новая рефлекторная дуга в процессе своего закладывания и дальнейшего закрепления»1 (подч. мной.— Л, £.).
ф Рецептивное поле доминанты создает спектр тех сигнально значимых факторов среды, которые обеспечивают данную доминанту на разных стадиях ее существования. А. А. Ухтомский (1966) выделил три стадии доминанты:
1. Стадия доминанты, безразлично привлекающей к себе в качестве поводов возбуждения всевозможные рецепции.
2. Стадия образования условного рефлекса, когда из множества действующих рецепций доминанта вылавливает группу рецепций, которая для нее в особенности биологически интересна (выработка адекватного раздражителя для данной доминанты). 3. Стадия более или менее исключительной связи данной доминанты с данным рецептивным содержанием, когда каждый из контрагентов (внутреннее содержание и внешний образ) будут вызывать и подкреплять исключительно друг друга.
На первой стадии достаточно устойчивой доминанты ее рецептивное поле оказывается чрезвычайно широким: разнообразные афферентации подкрепляют наличную доминанту. Причем уже на этой стадии обнаруживается координирующая роль торможения, препятствующего выполнению иных форм деятельности (Р. А. Павлыгина, 1982, 1988).
Признавая за доминантой способность подкрепляться разнообразными факторами среды, следует иметь в виду существование механизмов избирательного реагирования, т. е. наиболее эффективных агентов среды, жестко связанных с поддержанием и реализа-
*Ухтомский А. А. Собр. соч. Л., 1954. Т. IV. С. 225—226.
циеи данного доминантного состояния. К такого рода сигналам в первую очередь должны быть отнесены натуральные раздражители (по павловской терминологии) или ключевые раздражители (по терминологии этологов). Причем совсем не обязательно, чтобы такие свойства сигналов были бы генетически детерминированы.
Например, в определенных условиях одиночные коммуникационные сигналы животных могут приобретать характер ключевых раздражителей. Таким условием для кошек может быть возникновение биологической доминанты, проявляющейся во время лактации {Н. Н. Соколова, 1984). В стрессорных ситуациях котята издают сигналы, близкие по структуре к сигналам новорожденных. Такие сигналы неизменно вызывают реакцию материнского ухаживания у самок. Нсли же такие сигналы предъявляются с магнитофона, то у лактирующих самок проявляется лишь ориентировочная реакция без активного поиска котенка. Следовательно, для возникновения адекватных ответных реакций кошки-матери необходимо комплексное воздействие разномодального сигнала, звуковые же компоненты являются лишь частью демонстрационного поведенческого комплекса, недостаточной для полного воспроизведения адекватных реакций.
Таким образом, рецептивное поле доминанты материнского ухаживания у лактирующих кошек представляет собой динамическую систему, в состав которой входят жесткие компоненты (ключевые сигналы) и пластические компоненты, биологическая значимость которых для данной доминанты оценивается в контексте всей окружающей ситуации.
Фронтальная кора у кошек-самок к моменту рождения детенышей оказывается избирательно настроенной к тем звуковым частотам, которые являются формантными для видосиецифических вокализаций детенышей первого месяца жизни (Г. А. Куликов, 1989). Значит, состояние доминанты обеспечивает выделение жестких компонентов (ключевых), связанных с восприятием наи-
более значимой в биологическом отношении сигнализации и выполнением соответствующих жестко запрограммированных поведенческих актов.
Закономерности избирательного реагирования на сигнально значимые агенты среды могут служить физиологическими предпосылками для трактовки механизмов сенсорного внимания. В самом общем смысле внимание рассматривается как один из механизмов устранения избыточности сенсорных сообщений, который участвует как в избирательном подавлении, обеспечении сенсорных входов, так и 'в фильтрации информации, извлекаемой из систем кратко- и долговременной памяти.
Закрепление рецептивного поля с выделением адекватных для данной доминанты агентов внешней среды осуществляется по механизму условного рефлекса, в котором важнейшую роль отводят фактору подкрепления. Подкрепление условного рефлекса направлено к удовлетворению текущей доминирующей мотивации, к снижению возбудимости первичного доминантного очага. Мотивационное поведение животных направлено прежде всего на снижение доминанты. Полное удовлетворение биологической потребности, достигнутое в результате подкрепления систем условных рефлексов, приводит к исчезновению доминантных очагов и торможению соответствующих .условных рефлексов.
Теперь обратимся к рассмотрению эндогенно вызванного доминантного состояния (например, жажды) и его условно-рефлекторного удовлетворения (И. П. Никитина, 1983).
Мотивация жажды создавалась добавлением соли в пищевой рацион и ограничением воды. У животных на разные условные сигналы вырабатывались побежки к двум разным кормушкам, в одной из которых подавалась пресная, а в другой — соленая пища. При избыточном поступлении соли в организм возникал отказ животного даже от минимально соленой пиши и прекращение побежек по сигналу «соленого». В результате солевой перегрузки активируется система неспецифической
оценки содержания соли в предлагаемой животному пище с последующим формированием отрицательной реакции на эту пищу. Это состояние является стойким и гетеросенсорным по своему обеспечению, ибо не только конкретный условный сигнал, но и вся окружающая ситуация его подкрепляла, т. е. оно характеризовалось всеми свойствами доминантного состояния. Доминирующая мотивация жажды выступает как фактор неспецифической инициации реакций отвергания всего соленого и предпочтения всего пресного. Одновременно с этим могут возникнуть и реакции неспецифического поиска предмета удовлетворения доминирующей мотивации. Даже однократное положительное завершение такого поведенческого акта, санкционируемое соответствующим эмоциональным сопровождением, может закрепиться в памяти.
Значение сигнальных факторов окружающей среды выступает ярко при использовании различных приемов противопоставления условных сигналов «пресного», которые теперь подкрепляются пищей разной степени солености. По сигналам «пресного» животное направляется к соответствующей кормушке и поедает из нее пищу такой степени солености, которая ранее стойко отвергалась. Такое условно-рефлекторное «опреснение» пищи (психическое опреснение) может служить удачной моделью для исследования вкусовых иллюзий.
Вероятно, пищевой условный сигнал, как и доминирующая мотивация, управляют прежде всего вкусовыми порогами, поэтому животное и поедает неадекватную сигналу пищу. Условный сигнал приобретает свойства создавать определенный диапазон вкусовых порогов, в пределах которого данная мотивация может быть реализована с наибольшей вероятностью. Висцеральная сигнализация,"формирующая доминирующую мотивацию, также определяет тот пороговый диапазон вкусовых сигналов, которые способны удовлетворить биологическую потребность.
Третья стадия доминанты строится на законе «обратных отношений» А. А.
Ухтомского: симптомокомплекс доминанты может репродуцироваться при наличии адекватных сигналов. Такие ретенционные способности принадлежат коре больших полушарий, которая а) формирует адекватные стимулы для доминанты, б) запускает соответствующие доминанты по поводу этих стимулов и в) обеспечивает взаимодействие и переключение доминант.
На этой стадии речь должна идти не столько об элементарных биологических доминантах (голод, жажда, половое чувство), сколько о сложном памятном следе, носящем ситуативный характер (интегральный образ), а также о динамической констелляции мозговых структур, обеспечивающих целенаправленное поведение организма. Старая доминанта возобновляется или для того чтобы при новых данных обойтись при помощи старого опыта, или для того, чтобы по новым данным переинтегрировать старый опыт (А. А. Ухтомский, 1950). Значит, пространственно разнесенные компоненты доминирующей констелляции представляют собой центральную модель поведенческого акта, которая сформирована по закону доминанты и условного рефлекса и записана в долговременной памяти.
Справедливо заключение П. В. Симонова, который пишет: «Комбинация (точнее — синтез) механизма доминанты с механизмами формирования условного рефлекса обеспечивают оба фактора, необходимых и достаточных для организации целенаправленного поведения: его активный творческий характер (доминанта) и точное соответствие объективной реальности (упроченный, тонко специализированный условный рефлекс»)1.
Таким образом, доминанта и условный рефлекс рассматривается в качестве основных принципов организации целенаправленного и адаптивного поведения.
1 Симонов П. В. Взаимодействие доминанты и условного рефлекса как функциональная единица организации поведения//Утехи физиол. наук, 1983. Т. 14. № 1. С. 18—19.
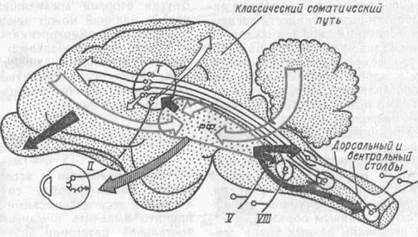
Рис. 68. Схема стволовойчасти мозга кошки, показывающая корковые связи
ретикулярной формации (по Дж. Френчу, 1962):
стрелками с точками показаны влияния на ретикулярную формацию (РФ); темные стрелки — тормозящие эффекты ствола мозга на сенсорную проводимость в мозгу; заштрихованные стрелки - облегчающие или тормозящие эффекты ствола мозга на рецептор; Т--таламус; II, V, VIII — черепно-мозговые нервы
Дата добавления: 2015-08-04; просмотров: 1810;