Понятие о моторной единице, ее свойства. Типы и режимы сокращения скелетных мыщц (изотоническое, изометрическое, одиночное, тетаническое).
Понятие о моторной единице. Ее св-ва.Моторная единица- это мотонейрон с группой иннервируемых им мышечных волокон. Кол-во мышечных волокон, входящих в моторную единицу различно (для глаза <10, пальцев рук 10-25, бицепсы 750). По скорости сокращения делятся на: -быстрые (фазные) например глазного яблока от 10-30мсек развивают большую силу при сокращении, неустойчивы к утомлению, иннервируются высокопороговыми нейронами, гладкий тетанус возникает при большей частоте стимуляции; -медленные тонические, длительность их сокращений >100мсек, развивают меньшую силу при сокращении, устойчивы к утомлению, иннервируются низкопороговыми нейронами, тетанус возникает при низкой частоте (мышцы спины). Типы и режимы сокращения скелетных мышц. Типы: -изотоническое (такое сокращение мышцы, при котором меняется длина, а напряжение постоянно (язык)); -изометрическое (когда оба конца мышцы неподвижно закреплены, длина не изменяется, повышается напряжение (тонус)); -ауксотоническое (при котором мышца развивает напряжение и укорачивается). Режим сокращений. Различают изотонический, изометрический и смешанный режимы сокращения мышц. При изотоническом сокращении мышцы происходит изменение ее длины, а напряжение остается постоянным. Такое сокращение происходит в том случае, когда мышца не перемещает груз. В естественных условиях близкими к изотоническому типу сокращений являются сокращения мышц языка. При изометрическом сокращении длина мышечных волокон остается постоянной, меняется напряжение мышцы. Такое сокращение мышцы можно получить при попытке поднять непосильный груз. В целом организме сокращения мышц никогда не бывают чисто изотоническим или изометрическим, они всегда имеют смешанный характер, т. е. происходит изменение и длины, и напряжения мышцы. Такой режим сокращения называется ауксотоническим если преобладает напряжение мышцы, или ауксометрическим если преобладает укорочение. Определяется частотой и пульсацией мотонейронов. Различают: одиночное мышечное сокращение и тетаническое. Одиночное это механический ответ мышечного волокна или отдельной мышцы на однократное их раздражение (пороговая или сверхпороговая сила)(сердечная мышца). Для скелетных мышц этот режим можно видеть только в лаборатории. Тетанус- это сильное длительное сокращение мышцы на ряд следующих друг за другом раздражителей при частоте >10им/сек. Различают: -зубчатый (возникает при малой частоте, когда последнее ее сокращение попадает на фазу расслабления, например дрожь); -гладкий (при большей частоте, когда последующий стимул приходится на фазу сокращения). Механизм мышечного сокращения и расслабления (теория скользящих нитей). Мышца состоит из мышечных волокон, которые состоят из тонких нитей миофибрилл (специализированный сократительный аппарат). Важнейшими компонентами ее являются: митохондрии, саркоплазматическая сеть (система продольных трубочек), Т-система (поперечных трубочек). Под световым микроскопом видно чередование темных и светлых дисков (поперечная исчерченность). Светлые диски- диски J- изотропные, не обладают двойным лучепреломлением. Темные диски- А диски- анизотропные, обладают двойным лучепреломлением. Функциональной единицей миофибрилла является саркомер (часть мышечного волокна от середины одного изотропного диска до середины другого. Z пластинки отделяют саркомеры друг от друга. Саркоплазматическим ретикулом является депо кальция (сокращение). Микрофибрилла состоит из белков актина и миозина. Миозиновые волокна толстые, короткие. Актиновые длинные, тонкие. На боковой стороне миозиновой нити имеются выступы под углом 120 к оси нити –поперечные мостики. Мостик состоит из головки, шейки и хвоста. Головка обладает АТФазными св-ми, шейками эластическими св-ми (за счет чего может поворачиваться вокруг своей оси). Актиновые компоненты имеют на своей поверхности активные центры, расположенные на расстоянии 40нм друг от друга, к ним прикрепляются головки миозина. Кроме актина в тонких нитях имеются и другие белки: тропонин и тропомиазин. Тропомиазин в покое блокирует актин. При возникновении нервного импульса потенциал действия распространяется по внешней мембране, затем по Т-системе. Начинается интенсивный выход кальция, достигая пороговой величины, затем взаимодействует с тропонином. При этом освобождает активные центры на актиновых нитях. Далее головка поперечного мостика присоединяется к активному центру актиновой нити, поворачивается вокруг своей оси на 45 и актиновые нити начинают скользить относительно миозиновых. При этом уменьшается длина мышцы и размер саркомера. Последовательная р-ция соединения и разъединения головок мостика ведет к скольжению нитей относительно друг друга по типу гребковых движений. Каждый мостик работает циклически, он тянет актин к центру А-полосы на расстояние 5-10нм, затем отщепляется от актина и присоединяется к нему в другой точке, более удаленной от центра А-полосы. На одно движение мостика тратиться одна молекула АТФ. При возбуждении АТФ распадается на АДФ и неорганический фосфат. С прекращением действия импульса кальций вновь поглащается Т-системой, отсоединяется от тропонина и тропомиазин вновь блокирует актин. Происходит распад мостиков и расслабление мышцы. Теплообразование: -теплота активации (быстрое выделение тепла на ранних этапах мышечного сокращения, когда отсутствуют признаки укорочения или развития напряжения); -теплота укорочения (выделение тепла при совершении работы); -теплота расслабления (при выделении тепла упругими элементами мышцы при расслаблении).
Механизмы мышечного сокращения и раслабления.Режимы мышечных сокращений. Различают изотонический, изометрический и смешанный режимы сокращения мышц. При изотоническом сокращении мышцы происходит изменение ее длины, а напряжение остается постоянным. Такое сокращение происходит в том случае, когда мышца не перемещает груз. В естественных условиях близкими к изотоническому типу сокращений являются сокращения мышц языка. При изометрическом сокращении длина мышечных волокон остается постоянной, меняется напряжение мышцы. Такое сокращение мышцы можно получить при попытке поднять непосильный груз. В целом организме сокращения мышц никогда не бывают чисто изотоническим или изометрическим, они всегда имеют смешанный характер, т. е. происходит изменение и длины, и напряжения мышцы. Такой режим сокращения называется ауксотоническим если преобладает напряжение мышцы, или ауксометрическим если преобладает укорочение.
Механизм мышечного сокращения. Мышцы состоят из мышечных волокон, которые состоят из множества тонких нитей - миофибрилл, расположенных продольно. Каждая миофибрилла состоит из протофибрилл - нитей сократительных белков актина и миозина. Перегородки, называемые 2-пластинами, разделяют миофибриллы и, следовательно, мышечное волокно на участки - саркомеры. В саркомере наблюдают правильно чередующиеся поперечные светлые и темные полосы. Эта поперечная исчерченность миофибрилл обусловлена определенным расположением нитей актина и миозина. В центральной части каждого саркомера свободно расположены толстые нити миозина. На обоих концах саркомера находятся тонкие нити актина, прикрепленные к Z-пластинам. Нити миозина выглядят в световом микроскопе как светлая полоска (Н-зона) в темном диске, который дает двойное лучепреломление, т. к. содержит нити миозина и актина и называется анизотропным или А-диском. По обестороны от А-диска находятся участки, которые содержат только тонкие нити актина и кажутся светлыми, т. к. они обладают одним лучепреломлением и называются изотропными или j-дисками. По их середине проходит темная линия - Z-мембрана. Именно благодаря такому периодическому чередованию светлых и темных дисков сердечная и скелетная мышцы выглядят исчерченными (поперечно-полосатыми) (рис. 6).
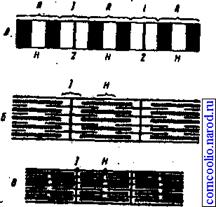
Рис. 6. Электронномикроскопическая картина миофибриллы (схематизировано)(А). Взаимное расположение толстых (миозиновых) и тонких (актиновых) нитей в расслабленной (Б) и сокращенной (В) миофибрилле. В состоянии покоя концы толстых и тонких: нитей лишь незначительно перекрываются на уровне А-диска. В соответствии с теорией скользящих нитей при сокращении тонкие актиновые нити скользят вдоль толстых миозиновых нитей, двигаясь между ними к середине саркомера. Сами актиновые и миозиновые нити своей длины не изменяют. Механизм скольжения нитей. Миозиновые нити имеют поперечные мостики (выступы) с головками, которые отходят от нити биполярно. Актиновая нить состоит из двух закрученных одна вокруг другой цепочек (подобно скрученным ниткам бус) молекул актина. На нитях актина расположены молекулы тропонина, а в желобках между двумя нитями актина лежат нити тропомиозина. Молекулы тропомиозина в покое располагаются так, что предотвращают прикрепление поперечных мостиков миозина к актиновым нитям. Во многих местах участки поверхностной мембраны мышечной клетки углубляются в виде трубочек внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-систему). Параллельно миофибриллам и перпендикулярно поперечным трубочкам расположена система продольных трубочек (альфа-система). Пузырьки на концах этих трубочек - терминальные цистерны - подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В этих пузырьках сосредоточено основное количество внутриклеточного кальция. В состоянии покоя миозиновый мостик заряжен энергией (миозин фосфорилирован), но он не может соединиться с нитью актина, так как между ними находится система из нитей тропомиозина и глобул тропонина. При возбуждении ПД быстро распространяется по мембранам поперечной системы внутрь клетки и вызывает высвобождение ионов кальция из альфа-системы. С появлением ионов кальция в присутствии АТФ происходит изменение пространственного положения тропонина, в результате чего отодвигается нить тропомиозина и открываются участки актина, присоединяющие ми-озиновые головки. Соединение головки фосфорилированного миозина с актином приводит к изменению положения мостика (его "сгибанию"), в результате конформации этой части миозиновой молекулы, и перемещению нити актина на один шаг (на один "гребок") к середине саркомера. Затем происходит отсоединение мостика от актина. Ритмические прикрепления и отсоединения головок миозина позволяют "грести" или тянуть актиновую нить к середине саркомера. При отсутствии повторного возбуждения ионы кальция закачиваются кальциевым насосом из протофибриллярного пространства в систему саркоплазматического ретикулума. Это приводит к снижению концентрации ионов кальция и отсоединению его от тропонина. Вследствие чего тропомиозин возвращается на прежнее место и снова блокирует активные центры актина. Вместе с тем, происходит фосфорилирование миозина за счет АТФ, который не только заряжает системы для дальнейшей работы, но и способствует временному разобщению нитей. Удлинение (расслабление) мышцы после ее сокращения является процессом пассивным, поскольку актиновые и миозиновые нити легко скользят в обратном направлении под влиянием сил упругости мышечных волокон и мышцы, а также силы растяжения мышц антагонистов.
11).Порог раздражения. Законы раздражения возбудимых тканей. Закон силы раздражения: чем сильнее раздражение, тем сильнее ответная реакция ткани до известного предела.
З. длительности раздражения: чем длиннее раздражение, тем сильнее ответная реакция организма до определённого предела.
З. «всё или ничего»: возбудимая ткань даёт максимальную ответную реакцию при любой силе порогового и непорогового раздр., и не даёт ничего при подпороговых стимулах.
12). Реобаза – минимальная сила постоянного тока, способная вызвать возбуждение(порог раздражения).
Хронаксия – время, в течение которого должен действовать ток удвоенной реобазы, чтобы вызвать возбуждение. Более возбудимая ткань имеет меньшую хронаксию.
Полезное время – наименьшее время, в течение которого должен действовать раздражающий стимул, величиной в одну реобазу. В клинике хронаксия – мерило возбудимости. Величина реобазы и хронаксии нервных волокон значительно меньше соответствующих величин мышечных волокон. Возбудимое образование может реагировать на стимул разной картиной ответа, поскольку для возбудимой ткани, клетки, органа или системы существует определённая физиологическая подвижность или лабильность.
Лабильность (функц. подвижность) – скорость элементарных реакций за единицу времени. Лабильность – максимальное число потенциалов действия, которое возбудимая ткань может генерировать в единицу времени в соответствии с частотой и ритмом раздражения. Наименьшая лабильность у синапсов, которые генерируют от80-120 имп/сек, для мышечного волокна в пределах 150-220имп/сек. Наиб. лабильностью обладает нервный проводник, например, аксоны типа А могут проводить до 3000имп/сек.
Оптимум – наивысшее сокращение мышцы происходит при нанесении на нерв нервно-мышечного препарата раздражений в более редком ритме. При этом ритме раздражений, каждый новый импульс возбуждения возникает во время фазы экзальтации, созданной предшествующим импульсом, в результате чего максимально сокращается мышца.
Пессимум – возникает при очень частых раздражениях, превышающих оптимальный ритм, сокращения мышцы уменьшаются и даже совсем прекращаются. Возникает, когда частота раздражений превышает меру лабильности. Под влиянием первого раздражителя ткань возбуждается, затем, когда импульс возбуждения ещё не закончился, наносится повторное раздражение – действует во время фазы абсолютной рефрактерности, она углубляется. Поэтому каждое новое раздражение посылаемое в очень частом ритме, снижает лабильность ткани и вызывает торможение.
14).Потенциал действия – быстрое колебание мембранного потенциала, возникающее при возбуждении. Потенциал действия – активный процесс, в основе которого лежит изменение ионной проницаемости мембраны. Потенциал действия регистрируется в нервных и мышечных клетках с помощью внутриклеточных электродов. Можно выделить следующие фазы развития ПД:1-деполяризации, продолжается около 0,2-0,5 мс.; 2 – Овершут; 3-реполяризации; 4-следовая деполяризация, при которой скорость реполяризации замедляется; 5-следовая гиперполяризация. Во время этой фазы мембрана становится более поляризованной, чем при потенциале покоя. Потенциал действия возникает при деполяризации мембраны примерно до -50мВ. Этот уровень называется пороговым. В начале действия раздражителя открывается лишь небольшая часть натриевых каналов. В результате происходит постепенная деполяризация мембраны, которая называется локальным потенциалом. Локальный переходит в потенциал действия при достижении порогового уровня. Лишь при этой деполяризации открывается такое количество натриевых каналов, которое приводит к развёртыванию всех фаз ПД. Изменения возбудимости носят следующий характер: при локальном ответе она повышается, в момент генерации ПД, Овершута отмечается её снижение (рефрактерность), во время реполяризации она вновь повышается. Во время следовой деполяризации отмечается повышенная возбудимость, следовательно гиперполяризация сопровождается снижение возбудимости. Ионные механизмы ПД: в основе возбуждения лежит повышение проводимости мембраны для натрия в 20 раз больше, чем для калия. Повышение проницаемости для калия – важный фактор в реполяризации мембраны. Итак, ПД обусловлен циклическим процессом входа натрия в клетку и последующего выхода калия. Причиной деполяризации, после генерации ПД является накопление ионов калия у наружной поверхности мембраны, что ведёт к увеличению мембранного потенциала. Следовая гиперполяризация связана преимущественно с активацией электрогенного насоса, вследствие накопления ионов натрия в клетке.
Рефрактерность (невосприимчивость) – кратковременное снижение возбудимости нервной и мышечной ткани во время и вслед за ПД. Рефрактерность это тоже фазный процесс. Абсолютный рефрактерный период – продолжается около 1 мс, состояние полной невозбудимоси, при действии раздражителя любой силы. Относительный рефрактерный период – когда путём значительной деполяризации можно вызвать ПД, хотя его амплитуда снижена по сравнению с нормой. Восстановление возбудимости происходит прежде всего в наиболее возбудимых волокнах. Возможность рефрактерности мембраны – важное следствие инактивации натриевой проницаемости.
15).Нервное волокно. Классификация. Механизм проведения возбуждения.
Различают безмиелиновые и миелиновые нерв. волокна. Безмиелиновые состоят из осевых цилиндров, окружённых шванновскими клетками. При погружении осевого цилиндра в швановскую клетку, её клеточная мембрана смыкается и образует мезаксон (сдвоенные мембраны шванновской клетки). Каждая ш. клетка подобным образом окружает несколько осевых цилиндров.
Миелиновое волокно состоит из осевого цилиндра, вокруг которого шванновские клетки образуют миелин за счёт концентрического наслаивания мембраны мезаксонов. Миелин прерывается через равные промежутки, которые называются перехватами Ранвье. Основной функцией нерва является проведение сигналов от рецепторов к нервному центру – аффирентные проводники, и от нервного центра на рабочий орган – эфферентные. Нервные проводники различаются диаметром, скорость проведения возобновляется, наличием или отсутствием миелина. 3 класса
А – толстые миелиновые волокна от 3-20мкн., v проведения волокон от 12-120м/с.
В – миелин. вол., преимущественно в вегетативной н.с., диаметром от 1-3 мкн., v от 3-14 м/с.
С – безмиелиновые – аффиренты рецепторов; эффиренты в.н.с., диаметром <1,5 мкн., v 0,5-2 м/с.
Безмиелиновые волокна на всём протяжении имеют одинаковую электрическую проводимость и сопротивление. В следствии d-поляризации участка мембраны, возникающий в нём локальный местный ток распространяется только на рядом расположенный не возб. участок – проведение возбуждения без декремента (остатка).
Миелиновое волокно имеет изолированный слой; перехваты, лишённые миелина имеют очень низкое сопротивление, поэтому являются центрами электрической активности. Путь передачи возбуждения – скачкообразный или сальтаторный. Преимущества миелиновых волокон:
возбуждение распространяется со значительно большей скоростью
энергетически более экономное, т.к. в состояние активности приходят небольшие участки мембраны в области перехватов.
невелика потеря ионов
малы затраты на работу натрий – калиевого насоса
быстро, точно и дифференцированно проводят различные виды чувствительности, обеспечивая максимально быстрые, адекватные реакции.
Законы проведения возбуждения по нервному волокну:
анатомич. и физ. целостности нерва – проведение импульсов возможно лишь при условии анатомической и физиологической целостности нервного проводника.
закон двустороннего проведения возбуждения по нерву. При раздражении нервного волокна возбуждение распространяется по нему и в центробежном и в центростремительном направлении. В целостном организме данный закон не действует, т.к. имеет место одностороннее проведение возбуждения, за счёт химических синапсов.
закон изолированного проведения возбуждения по нерву, применим только для миелиновых волокон. Говорит о том, что в периферическом нерве импульсы распространяются изолированно по каждому волокну, не переходя с одного волокна на другое.
Основные механизмы проводниковой блокады. Обязательным условием проведения возбуждения по нерву является анатомическая и функциональная целостность мембраны осевого цилиндра. Разные физические воздействия (перевязка, охлаждение, согревание), а также хим. вещества (новокаин, хлороформ, дикаин) нарушают генерацию импульсов, прекращают проведение из по нерву. При нарушении физиологической целостности нервного волокна происходит блокада натриевых каналов возбудимой мембраны, торможение волны деполяризации.
16).Синапс – специализированная структура, обеспечивающая передачу возбуждения с нервного волокна на эффекторную клетку – мышечную, нервную или секреторную, или развитие торможения в этих структурах.
Синапсы подразделяются на : аксо-соматические, аксо-аксональные, аксо-дендритические, дендро-соматические, дендро-дендритические, нервно-мышечные, и т.д. Все синапсы делятся на центральные (в головном и спинном мозге) и периферические (нервно-мышечные, синапсы вегетативных ганглиев).
По виду медиатора: адренергические и холинергические. По способу передачи возбуждения: химические, электрические и смешанные. По конечному физиологическому эффекту на возбуждающие и тормозные.
Синапс как объект воздействия и точка приложения лекарственных средств. Лекарственные препараты могут влиять на различные этапы синаптической передачи, вызывая положительные или отрицательные реакции организмов и систем. Например миорелаксан. Блокируют передачу возбуждения в нервно-мышечных синапсах. Блокада нервно-мышечной передачи может быть вызвана местно-анестезирующими веществами, например: новокаин – блокирует возбуждение пресинаптической части. Ботулинический токсин блокирует выделение медиатора. Яд кураре вытесняет ацетилхолин из рецепторов. Действие многих лекарственных веществ имеет точкой приложения мозг и направлено на прерывание или облегчение синаптической передачи. Психотропные вещества: нейролептики – стимулируют выделение медиаторов, транквилизаторов; седативные препараты ослабляют выделение медиатора. Многие препараты связываются с рецепторами головного мозга и блокируют их.
19).Моторная единица – мота нейрон с группой иннервируемых или мышечных волокон. Количество мышечных волокон, входящих в моторную единицу различно. Для пальцев руки 10-35. По скорости сокращения:
быстрые или фазные()гл.яблоко
, длит. сокр. 10-30 м/с. Развивают большую силу при сокращении, менее устойчивы к утомлению, иннервируются высокопороговыми нейронами. Гладкий тетанус – возникает при большей частоте стимуляции.
медл.томич.двиг.ед., длительность сокращения – 100 м/с. Развивают меньшую силу при сокращении, более устойчивы к утомлению. Иннервируются низкопороговыми нейронами. Тетанус возникает при низкой частоте (мышцы спины).
Типы и режимы сокращения скелетных мышц.
изотоническое – такое сокращение, при котором меняется длина мышцы (её волокна укорачиваются), но напряжение остаётся постоянным.
изометрическое – оба конца мышцы неподвижно закреплены, длина мышцы не изменяется, а значительно повышается тонус(напряжение мышцы).
одиночное – механический ответ мышечного волокна или отдельной мышцы на однократное их раздражение. Этот режим можно наблюдать для скелетных мышц только в лабораторных условиях. В организме лишь сердце работает в таком режиме. ОМС состоит из нескольких периодов:
латентный – с момента нанесения раздражения до видимого сокращения (0,01с).
Период сокращения, восходящей энергии (0,03-0,04 с).
Период расслабления, нисходящей энергии (0,04-0,06 с).
4.тетаническое – сильное и длительное мышечное сокращение на ряд следующих друг за другом раздражителей. Различают гладкий тетанус – при большой частоте, когда последующий стимул приходится в фазу сокращения. Зубчатый – при относительно малой частоте, когда последнее сокращение попадает в фазу расслабления и амплитуда второго сокращения выше, чем предыдущего.
Дата добавления: 2015-08-01; просмотров: 5577;