Основные физиологические состояния биологической мембраны. Потенциал покоя. Мембранно-ионная теория происхождения потенциала покаоя. Натрий-калиевый насос.
Мембранный 'потенциал (МП) - 'потенциал покоя, это трансмембранная разность потенциалов существующая между цитоплазмой и окружающей клетку средой. В состоянии покоя внутренний потенциал клетки отрицателен по отношению к наружному, условно принимаемому за 0. МП варьирует от -50 до - 90 мВ Для измерения ИП применяют микроэлектродную технику. Потенциал покоя изменяется при приложении электрического тока, изменении ионного состава среды, воздействии токсинов, недостатке кислорода Ксндл внутренний потенциал уменьшается (становится менее отрицательным) говорят о деполризации мембраны Увеличение отрицательного заряда внутри клетки называют гиперполяризацией.
Гснсз потенциала покоя» Наличие электрических потенциалов обусловлено неравенством концентрации конов натрия, калия, кальция и хлора внутри и вне клетки. А также различной проницаемостью для них поверхностной мембраны. Концентрация калия в цитоплазме нервных и мышечных клеток в 40-50 раз больше, чем вне клетки. Концентрационный градиент калия является основным фактором, определяющим величину ПП нервного волокна. Однако в покое мембрана проницаема и для натрия, диффузия которого во внутрь клетки уменьшает отрицательный потенциал клетки Ионы хлора в нервных волокнах не играют существенной роли в генезе ПП, проницаемость для них мембраны относительно мала Натрий-калиевый насос обеспечивает выведение из цитоплазмы, проникающих в него натрия и введение в цитоплазму калия; на,сос перемещает натрий и калий против их концентрационного градиента. Источник энергии для этой работы АТФ.
Электрофизиологическая характеристика потенциала действия, его фазы (деполяризация, реполяризация, отрицательный и положительный следовой потенциал). Изменение возбудимости мембранный в различные фазы одиночного цикла возбуждения. Понятие о рефрактерности.
Потенциал действия - быстрое колебание мембранного потенциала, возникающее при возбуждений. Потенциал действия - это активный процесс в основе которого лежит изменение ионной "проницаемости мембраны. Потенциал действия регистрируется в нервных и мышечных клетках с помощью внутриклеточных электродов. Можно четко выделить следующие фазы развития ИД: 1). Фаза деполяризации, продолжается около 0,2-0,5 мс; 2) Овершут; 3). Фаза реполярвзации.; 4). Следовая деполяризация, при которой скорость реполяризадии замедляется; 5). Следовал гиперполяризация. Во время этой фазы мембрана становится более поляризованной, чем при потенциале покоя, Потенциал действий возникает при деполяризации мембраны примерно до -50 мВ, Этот уровень именуется пороговым, В начале действия раздражителя открывается лишь небольшая часть "натриевых^каналов. В результате происходит постепенная деполяризация мембраны, которая именуется локальным (местным) потенциалом. Местный потенциал, переходит в потенциал действия при достижении порогового уровня, Лишь при этой деполяризации открывается такое количество натриевых каналов, которое приводит к развертыванию всех фаз ПД.
Изменения возбудимости носят следующий характер; при локальном ответе она повышается, в момент генерации ПД, овершута отмечается ее снижение (рефрактерность), во время реполяризации она вновь повышается. Во время следовой деполяризации отмечается повышенная возбудимость, следовая гиперполяризация сопровождается снижением возбудимости. Ионные механизмы ПД: в основе возбуждения лежит повышение проводимости мембраны для натрия в 20 раз больше, чем для калия. Повышение проницаемости для калия - важный фактор реполяризации мембраны. Итак, ПД обусловлен циклическим процессом входа натрия в клетку и последующего выхода калия. Причиной деполяризации, после генерации ПД является накопление ионов калия у наружной поверхности мембраны, что ведет к увеличению МП (следовой деполяризационный потенциал). Следовая гиперполяризация связана преимущественно с активацией электрогенного натриевого насоса, вследствие накопления ионов натрия в клетке. Рефрактерность (невосприимчивость) — кратковременное снижение возбуждения нервной и мышечной ткани во время и в след за ПД. Рефрактерность это тоже фазный процесс. Абсолютный рефрактерный период ~ продолжается около 1 мс, состояние полной невозбудимости, при действии раздражителя любой силы Относительный рефрактерный период - когда путем значительной деполяризации можно вызвагь ПД, хотя его амплитуда снижена по сравнению с нормой. Востановление возбудимости происходит прежде всего в наиболее возбудимых волокнах. Возможность рефрактерности мембраны — важное следствие инактивации натриевой проницаемости.
Нервное волокно. Классификация и морфофизиологическая характеристика нервных волокон. Механизм проведения возбуждения в миелиновых и безмиелиновых нервных волокнах. Законы проведения возбуждения по нервному волокну. Основные механизмы фармакологической блокады проведения возбуждения.
Нервные волокна (отростки нервных клеток) выполняют специализированную функцию - проведение нервных импульсов. По морфологическому признаку нервные волокна делятся на миелиновые (покрытые миелиновой оболочкой) и безмиелиновые. Нервные волокна формируют нерв или нервный ствол, состоящий из большого числа нервных волокон, заключенных в общую соединительноткан-ную оболочку. В состав нерва входят миелиновые и безмиелиновые волокна. Нервные волокна, проводящие возбуждение от рецепторов в ЦНС называются афферентными, а волокна, проводящие возбуждение от ЦНС к исполнительным органам, называются эфферентными. Нервы состоят из афферентных и эфферентных волокон. Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью. Проведение возбуждения по нервным волокнам осуществляется по определенным законам. Закон двустороннего проведения возбуждения по нервному волокну. Возбуждение по нервному волокну распространяется в обе стороны от места его возникновения, т. е. центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Закон анатомической и физиологической целостности нервного волокна. Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность. Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е. к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, т. е. не переходя с одного волокна на другое. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна.
Изолированное проведение возбуждения имеет важное значение. Нервсодержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки, ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно. Нервные волокна по скорости проведения возбуждения, диаметру волок- на, длительности различных фаз потенциала действия и строению принято подразделять на три типа: А, В, С. Волокна типа А в свою очередь делятся на подтипы: А-альфа, А-бета, А-гамма, А-дельта. Волокна типа А покрыты миелиновой оболочкой. Наиболее толстые из них А - альфа, они имеют диаметр 12-22 мкм и высокую скорость проведения возбуждения - 70-120 м/с. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от определенных рецепторов мышц к соответствующим нервным центрам. Три другие группы волокон тина А (бета, гамма, дельта) имеют меньший диаметр от 8 до 1 мкм и меньшую скорость проведения возбуждения от 5 до 70 м/с. Волокна этих групп относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых, рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к интрафузальным мышечным волокнам. К волокнам типа В относятся миелинизированные преганглио-нарные волокна вегетативной нервной системы. Их диаметр - 1-3,5 мкм, а скорость проведения возбуждения - 3-18 м/с.
К волокнам типа С относятся безмиелиновые нервные волокна малого диаметра - 0,5-2,0 мкм. Скорость проведения возбуждения в этих волокнах не более 3 м/с (0,5-3,0 м/с) . Большинство волокон типа С - это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления. Механизм проведения возбуждения по нервным волокнам. Механизм распространения возбуждения у различных нервных волокон неодинаков. По современным представлениям распространение возбуждения по нервным волокнам осуществляется на основе ионных механизмов генерации потенциала действия. При распространении возбуждения по безмиелиновому нервному волокну местные электрические токи,, которые возникают между его возбужденным участком, заряженным отрицательно, и невозбужденным, заряженным положительно, вызывают деполяризацию мембраны до критического уровня с последующей генерацией ПД в ближайшей точке невозбужденного участка мембраны. Этот процесс повторяется многократно. На всем протяжении нервного волокна происходит процесс репродукции нового ПД в каждой точке мембраны волокна. Такое проведение возбуждения называете и непрерывным. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки (перехватов Ранвье) создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. Местные электрические токи возникают между соседними перехватами Ранвье, т. к. мембрана возбужденного перехвата становится заряженной отрицательно по отношению к поверхности соседнего невозбужденного перехвата. Эти местные токи деполярязуют мембрану невозбужденного перехвата до критического уровня и в нем возникает ПД (рис. 4). Следовательно, возбуждение как бы "перепрыгивает" через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Такой механизм распространения возбуждения называется сальтаторным или скачкообразным. Скорость такого способа проведения возбуждения значительно выше и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов.
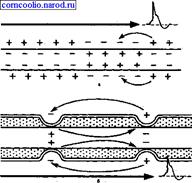
Рис. 4. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах.
"Перепрыгивание" потенциала действия через участок между перехватами возможно потому, что амплитуда ПД в 5-6 раз превышает пороговую величину, необходимую для возбуждения соседнего перехвата. ПД может "перепрыгивать" не только через один, но и через два межперехватных промежутка. Это явление может наблюдаться при снижении возбудимости соседнего перехвата под действием какого-либо фармакологического вещества, например, новокаина, кокаина и др. Нервные волокна обладают лабильностью - способностью воспроизводить определенное количество циклов возбуждения в единицу времени в соответствии с ритмом действующих раздражителей. Мерой лабильности является максимальное количество циклов возбуждения, которое способно воспроизвести нервное волокно в единицу времени без трансформации ритма раздражения. Лабильность определяется длительностью пика потенциала действия, т. е. фазой абсолютной рефрактерности. Так как длительность абсолютной ре-фрактерности у спайкового потенциала нервного волокна самая короткая, то лабильность его самая высокая. Нервное волокно способно воспроизвести до 1000 импульсов в секунду. Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента) посредством, например, отравления или повреждения, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности было названо Н. Е. Введенским парабиозом. В развитии состояния парабиоза можно отметить три, последовательно сменяющих друг друга, фазы: уравнительную, парадоксальную, тормозную. В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители - больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т. е. 25 импульсов из 100) , то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц) - происходит уравнивание ответной реакции. В парадоксальную-фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно, наблюдается парадокс - на редкие раздражители ответная реакция больше, чем на частые. В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние. Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ также связано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам.
Парабиоз - явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности. Механизм развития парабиотического состояния сводится к следующему. При воздействии на нервное волокно парабиотического фактора нарушается способность мембраны увеличивать натриевую проницаемость в ответ на раздражение. В участке альтерации инак-тивация натриевых каналов, вызванная повреждающим агентом, суммируется с инактивацией, вызываемой нервным импульсом, и возбудимость снижается настолько, что проведение следующего импульса блокируется.
Определение понятия «синапс». Строение синапса, классификация, свойства. Механизм синаптической передачи. Синапс как объект воздействия и приложения лекарственных веществ, ядов и токсинов.
Синапс - это специализированная структура, которая обеспечивает передачу возбуждения с одной возбудимой структуры на другую. Термин "синапс" введен Ч. Шеррингтоном и означает "сведение", "соединение", "застежка". Классификация синапсов. Синапсы можно классифицировать по:
1) их местоположению и принадлежности соответствующим структурам:
• периферические (нервно-мышечные, нейро-секреторные, рецеп-торнонейрональные);
• центральные (аксо-соматические, аксо-дендритные, аксо-аксо-нальные, сомато-девдритные, сомато-соматические);
2) знаку их действия - возбуждающие и тормозящие;
3) способу передачи сигналов - химические, электрические, смешанные.
4) медиатору, с помощью которого осуществляется передача - холинергические, адренергические, серотонинергические, глицинергические и т. д.
Строение синапса. Все синапсы имеют много общего, поэтому строение синапса и механизм передачи возбуждения в нем можно рассмотреть на примере нервно-мышечного синапса (рис. 7).
Синапс состоит из трех основных элементов:
• пресинаптической мембраны (в нервно-мышечном синапсе - это утолщенная концевая пластинка);
• постсинаптической мембраны;
• синаптической щели.
Пресинаптическая мембрана - это часть мембраны нервного окончания в области контакта его с мышечным волокном. Постси-наптическая мембрана - часть мембраны мышечного волокна. Часть постсинаптической мембраны, которая расположена напротив пресинаптической, называется субсинаптической мембраной. Осо-бенностью субсинаптической мембраны является наличие в ней специальных рецепторов, чувствительных к определенному медиатору, и наличие хемозависимых каналов. В постсинаптической мем-брне, за пределами субсинаптической, имеются потенциалозависимые каналы.
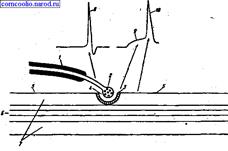
Рис. 7. Строение синапса (схема). 1 - миелинизированное нервное волокно; 2 - нервное окончание с пузырьками медиатора; 3 - субсинаптическая мембрана мышечного волокна; 4 - синаптическая щель; 5 - постсинаптическая мембрана мышечного волокна; 6 - миофибриллы; 7 - саркоплазма; 8 - потенциал действия нервного волокна; 9 - потенциал концевой пластинки (ВПСП); 10 - потенциал действия мышечного волокна.
Механизм передачи возбуждения в химических возбуждающих синапсах. В синапсах с химической передачей возбуждение передатся с помощью медиаторов (посредников). Медиаторы - это химические вещества, которые обеспечивают передачу возбуждения в синапсах. Медиаторы в зависимости от их природы делятся на несколько групп:
• моноамины (ацетилхолин, дофамин, норадреналин, серотонин и др.);
• Аминокислоты (гамма-аминомасляная кислота - ГАМК, глутаминовая кислота, глицин и др.);
• нейропептиды (вещество Р, эндорфины, нейротензин, АКТГ, ангиотензин, вазопрессин, соматостатин и др.). Медиатор в молекулярном виде находится в пузырьках пресинаптического утолщения (синаптической бляшке), куда он поступает:
• из околоядерной области нейрона с помощью быстрого аксо-нального транспорта (аксотока);
• за счет синтеза медиатора, протекающего в синаптических терминалях из продуктов его расщепления;
• за счет обратного захвата медиатора из синаптической щели в неизменном виде.
Когда по аксону к его терминалям приходит возбуждение, пресинаптическая мембрана деполяризуется, что сопровождается поступлением ионов кальция из внеклеточной жидкости внутрь нервного окончания. Поступившие ионы кальция активируют перемещение синаптических пузырьков к пресинаптической мембране, их соприкосновение и разрушение (лизис) их мембран с выходом медиатора в синаптическую щель. В ней медиатор диффундирует к суб-синаптической мембране, на которой находятся его рецепторы. Взаимодействие медиатора с рецепторами приводит к открытию преимущественно каналов для ионов натрия. Это приводит к деполяризации субсинаптической мембраны и возникновению так называемого возбуждающего постсинаптического потенциала (ВПСП). В нервно-мышечном синапсе ВПСП называется потенциалом концевой пластинки (ПКП). Между деполяризованной субсинаптической мембраной и соседними с ней участками постсинаптической мембраны возникают местные токи, которые деполяризуют мембрану. Когда они деполяризуют мембрану до критического уровня, в постсинаптической мембране мышечного волокна возникает потенциал действия, который распространяется по мембранам мышечного волокна и вызывает его сокращение. Химические тормозные синапсы. Эти синапсы по механизму передачи возбуждения сходны с синапсами возбуждающего действия. тормозных синапсах медиатор (например, глицин) взаимодействует с рецепторами субсинаптической мембраны и открывает в ней хлорные каналы, это приводит к движению ионов хлора по концентрационному градиенту внутрь клетки и развитию гиперполяризации на субсинаптической: мембране. Возникает так называемый тормозной постсинаптический потенциал (ТПСП). Ранее полагали, что каждому медиатору соответствует специфическая реакция постсинаптической клетки - возбуждение или торможение в той или иной форме. В настоящее время установлено, что одному медиатору чаще всего соответствует не один, а несколько различных рецепторов. Например, ацетилхолин в нервно-мышечных синапсах скелетных мышц действует на Н-холинорецепторы (чувствительные к никотину), которые открывают широкие каналы для натрия (и калия), что порождает ВПСП (ПКП) В ваго-сердечных синапсах тот же ацетилхолин действует на М-холинорецепторы (чувствительные к мускарину), открывающие селективные каналы для ионов калия, поэтому здесь генерируется тормозной постсинаптический потенциал (ТПСП). Следовательно, возбуждающий или тормозной характер действия медиатора определяется свойствами субсинаптической мембраны (точнее, видом рецептора), а не самого медиатора. Физиологические свойства химических синапсов. Синапсы с химической передачей возбуждения обладают рядом общих свойств:
• Возбуждение через синапсы проводится только в одном направлении (односторонне). Это обусловлено строением синапса: медиатор выделяется только из пресинаптического утолщения и взаимодействует с рецепторами субсинаптической мембраны;
• передача возбуждения через синапсы осуществляется медленнее, чем по нервному волокну - синаптическая задержка;
• передача возбуждения осуществляется с помощью специальных химических посредников - медиаторов;
• в синапсах происходит трансформация ритма возбуждения;
• синапсы обладают низкой лабильностью;
• синапсы обладают высокой утомляемостью;
• синапсы обладают высокой чувствительностью к химическим (в том числе и к фармакологическим) веществам. Электрические синапсы возбуждающего действия. Кроме синапсов с химической передачей возбуждения преимущественно в центральной нервной системе (ЦНС) встречаются синапсы с электрической передачей. Возбуждающим электрическим синапсам свойственны очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран, что обеспечивает эффективное прохождение локальных электрических токов. Низкое сопротивление, как правило, связано с наличием поперечных каналов, пересекающих обе мембраны, т. е. идущих из клетки в клетку (щелевой контакт). Каналы образуются белковыми молекулами (полумолекулами) каждой из контактирующих мембран, которые соединяются комплементарно. Эта структура легко проходима для электрического тока. Схема передачи возбуждения в электрическом синапсе: ток, вызванный пресинаптическим потенциалом действия, раздражает по-стсинаптическую мембрану, где возникает ВПСП и потенциал действия. Поперечные каналы объединяют клетки не только электрически, но и химически, так как они проходимы для многих низкомолекулярных соединений. Поэтому возбуждающие электрические синапсы с поперечными каналами формируются, как правило, между клетками одного типа (например, между клетками сердечной мышцы). Общими свойствами возбуждающих электрических синапсов являются:
• быстродействие (значительно превосходит таковое в химических синапсах);
• слабость следовых эффектов при передаче возбуждения (в результате этого в них практически невозможна суммация последовательных сигналов);
• высокая надежность передачи возбуждения. Возбуждающие электрические синапсы могут возникать при благоприятных условиях и исчезать при неблагоприятных. Например, при повреждении одной из контактирующих клеток ее электрические синапсы с другими клетками ликвидируются. Это свойство называется пластичностью. Электрические синапсы могут быть с односторонней и двусторонней передачей возбуждения. Электрический тормозный синапс. Наряду с электрическими синапсами возбуждающего действия могут встречаться электрические тормозные синапсы. Примером такого синапса может служить синапс, который образует нервное окончание на выходном сегменте маутнеровского нейрона у рыб. Тормозящее влияние возникает за счет действия тока, вызванного потенциалом действия пресинапти-ческой мембраны. Пресинаптический потенциал вызывает значительную гиперполяризацию сегмента и гиперполяризующий ток мгновенно тормозит генерацию потенциала действия в начальном сегменте аксона. В смешанных синапсах Пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Мышечная ткань, классификация, ультраструктура, функции.У человека различают три вида мышц:• поперечно-полосатые скелетные мышцы;• поперечно-полосатая сердечная мышца;• гладкие мышцы внутренних органов, кожи, сосудов.Мышцы обладают физическими и физиологическими свойствами. Рассмотрим те свойства, которые характерны для скилетных мышц.Физические свойства скелетных мышц.1. Растяжимость - способность мышцы изменять свою длину под действием растягивающей ее силы. 2. Эластичность - способность мышцы принимать свою первоначальную длину после прекращения действия растягивающей или деформирующей силы. Живая мышца обладает малой, но совершенной эластичностью: уже небольшая сила способна вызвать относительно большое удлинение мышцы, а возвращение ее к первоначальным размерам является полным. Эти свойства очень важны для осуществления нормальных функций скелетных мышц. 3. Сила мышцы. Она определяется максимальным грузом, который мышца в состоянии поднять. Для сравнения силы различных мышц определяют их удельную силу - максимальный груз, который мышца в состоянии поднять, делят на число квадратных сантиметров ее физиологического поперечного сечения. 4. Способность мышцы совершать работу. Работа мышцы определяется произведением величины поднятого груза на высоту подъема. Работа мышцы постепенно увеличивается с увеличением груза, но до определенного предела, после которого увеличение груза приводит к уменьшению работы, т. к. снижается высота подъема груза. Следовательно, максимальная работа мышцей производится при средних величинах нагрузок (закон средних нагрузок). Физиологические свойства мышц.
• Возбудимость - способность приходить в состояние возбуждения при действии раздражителей. • Проводимость - способность проводить возбуждение. • Сократимость - способность мышцы изменять свою длину или напряжение в ответ на действие раздражителя. • Лабильность - лабильность мышцы равна 200-300 Гц. При непосредственном раздражении мышцы (прямое раздражение) или опосредовано через иннервирующий ее двигательный нерв (непрямое раздражение) одиночным стимулом возникает одиночное мышечное сокращение, в котором выделяют три фазы:
• латентный период - время от начала действия раздражителя до начала ответной реакции; • фаза сокращения (фаза укорочения); • фаза расслабления. В естественных условиях к скелетной мышце из ЦНС поступают не одиночные импульсы, а серия импульсов, следующих друг за другом с определенными интервалами, на которую мышца отвечает длительным сокращением. Такое длительное сокращение мышцы, возникающее в ответ на ритмическое раздражение получило название тетанического сокращения или тетануса. Различают два вида тетануса: зубчатый и гладкий. Если каждый последующий импульс возбуждения поступает к мышце в тот период, когда она находится в фазе укорочения, то возникает гладкий тетанус, а если в фазу расслабления - зубчатый тетанус.
Амплитуда тетанического сокращения превышает амплитуду одиночного мышечного сокращения. Исходя из этого Гельмгольц объяснил процесс тетанического сокращения простой суперпозицией, т. е. простой суммацией амплитуды одного мышечного сокращения с амплитудой другого. Однако в дальнейшем было показано, что при тетанусе имеет место не простое сложение двух механических эффектов, т. к. эта сумма может быть то большей, то меньшей. Н. Е. Введенский объяснил это явление с точки зрения состояния возбудимости мышцы, введя понятие об оптимуме и пессимуме частоты раздражения. Оптимальной называется такая частота раздражения, при которой каждое последующее раздражение осуществляется в фазу повышенной возбудимости. Тетанус при этом будет максимальным по амплитуде – оптимальным. Пессимальной называется такая частота раздражения, при которой каждое последующее раздражение осуществляется в фазу пониженной возбудимости. Тетанус при этом будет минимальным по амплитуде – пессимальным.
Дата добавления: 2015-08-01; просмотров: 4262;