Лекция 3. Молекулярные основы наследственности, методы генной инженерии, генетически модифицированные организмы. – 2 часа
Молекулярная генетика исследует процессы, связанные с наследственностью на молекулярном уровне. Носителем наследственной информации является ДНК (у ретровирусов –РНК), а ген – это участок молекулы ДНК, ответственный за формирование какого- либо признака. ДНК и РНК – биополимеры, мономерами которых являются нуклеотиды. Нуклеотид состоит из трех химических веществ: углевода(У), остатка фосфорной кислоты (Ф) и азотистого основания (А):
Ф-У-А.
ДНК отличается от РНК углеводом: ДНК содержит дезоксирибозу, а РНК 0 рибозу.
В ДНК содержатся азотистые основания: аденин, гуанин, Тимин и цитозин. В молекуле РНК Тимин заменяется на на урацилю Молекула, как правило, двухцепочечная, а РНК – одноцепочечная.
Нуклеотиды двух цепочек соединяются с помощью азотистых оснований с помощью азотистых оснований по правилу комплементарности (дополнительности): аденин с Тимином, а гуанин с цитозином.
В раскрученном виде ДНК имеет подобие лестницы. Ее прочные ковалентные связи – перила образованы сахсрами и фосфатами, а ступени – азотистыми основаниями, между которыми имеются более слабые водородные связи:
Ф-Д-Ф-Д-Ф-Д-Ф-Д-Ф-Д
I I I I I
A Г Ц Т Г
: : : : :
Т Ц Г А Ц
I I I I I
Ф-Д-Ф-Д-Ф-Д-Ф-Д-Ф-Д
Последовательность расположения нуклеотидных пар создает специфичность молекулы ДНК и позволяет (при огромной ее длине) зашифровать большое количество информации.
ДНК способна к авторепродукции, т.е. самовоспроизведению. Процесс этот происходит в период S-интерфазы перед митозом или мейозом. Перед делением клетки на две дочерние все молекулы ДНК должны воспроизвести подобные им молекулы для того, чтобы каждая дочерняя клетка получила полный набор молекул ДНК и полный объем генетической информации, содержащейся в ДНК материнской клетки. При авторепродукции каждая молекула ДНК распадается на две одиночные цепи (при помощи фермента ДНК- нуклеазы), а затем на каждой материнской цепи достраивается дочерняя из свободных нуклеотидов по правилу комплементарности (при помощи фермента ДНК-полимеразы).
Реализация генетической информации происходит в процессе биосинтеза белков. Белки- органические соединения, играющие важную роль в жизнедеятельности живых организмов, 80% которых составляют клеточные структуры; белковую природу имеют почти все ферменты—катализаторы биохимических реакций, многие гормоны. Сократительные белки мышц обуславливают движение, а белки иммуноглобулины – защиту организма от инфекций.
Несмотря на многообразие белков, их видовую и тканевую специфичность молекулы имеют единый план строения. В состав белковой молекулы могут входить до 20 различных аминокислот. Аминокислоты соединяются в цепь, образуя полипептид. Молекула белка представляет собой одну или несколько связанных между собой полипептидных цепей. Первичная структура белка зашифрована на структурных генах молекулы ДНК (или РНК). Одну аминокислоту шифрует один триплет (кодон) – три расположенных рядом в цепочке ДНК (или РНК) нуклеотида.Из четырех нуклеотидов нуклеиновых кислот можно составить 64 триплета. Из них 61 являются смысловыми (то есть шифруют какую-то аминокислоту) и 3 – бессмысленными (нонсенсы), так называемые стоп-кодоны. Последние не кодируют аминокислот, а означают окончание информации о строении молекулы белка.
Синтез белка осуществляется в два этапа: транскрипции и трансляции.
1. Транскрипция – это переписывание генетической информации с молекулы ДНК на молекулу и-РНК (информационную, или матричную, РНК). Днк деспирализуется, рвутся водородные связи между азотистыми основаниями, и цепочки расходятся на участке какого-либо гена. Затем на смысловой цепочке ДНК, на участке структурного гена, синтезируется молекула и- РНК ( при помощи фермента РНК- полимеразы) по правилу комплементарности (А-У; Г-Ц) из свободных нуклеотидов.
2. . Трансляция – этап синтеза белка, происходящий в рибосомах ядра и цитоплазмы. В нем активное участие принимают транспортные РНК (т-РНК), которые подносят к рибосоме активированные аминокислоты и участвуют в расшифровке генетического кода. Небольшие молекулы (70-90 нуклеотидов) т-РНК способны сворачиваться таким образом, что образуют структуру, напоминающие по форме клеверный лист. В клетке имеется столько же разных т- РНК, сколько кодонов, шифрующих аминокислоты. На вершине среднего «листа» имеется распознающий триплет- антикодон.
3. Последовательность триплетов в молекуле и- РНК определяет последовательность аминокислотных остатков в синтезируемой на данной матрице молекуле белка..
УУГ УЦУ АЦУ и-РНК
Лейцин серин треонин белок
Только в том случае, если антикодон т-РНК комплементарен кодону и-РНК, т-РНК оставляет в рибосоме принесенную аминокислоту, и она присоединяется к синтезируемой белковой цепочке.
В рибосоме могут находиться одновременно две т-РНК. Таким образом, зная строение ДНК (гена), можно расшифровать структуру молекулы белка. Например:
ДНК: ААЦТГАЦАЦ
и-РНК: УУГАЦУГУГ
белок: лей-тре-вал.
Если известны изменения в ДНК, то можно предвидеть изменения в структуре белка.
Возможности генной инженерии.Генетическая инженерия родившись в начале 70-х годов,
Методы генетической инженерии.Генетическая инженерия - конструирование in vitro функционально активных генетических структур (рекомбинантных ДНК), или иначе - создание искусственных генетических программ (Баев А. А.). По Э. С. Пирузян генетическая инженерия - система экспериментальных приемов, позволяющих конструировать лабораторным путем (в пробирке) искусственные генетические структуры в виде так называемых рекомбинантных или гибридных молекул ДНК.
Речь идет о направленном, по заранее заданной программе конструировании молекулярных генетических систем вне организма с последующим введением их в живой организм. При этом рекомбинантные ДНК становятся составной частью генетического аппарата рецепиентного организма и сообщают ему новые уникальные генетические, биохимические, а затем и физиологические свойства.
Цель прикладной генетической инженерии заключается в конструировании таких рекомбинантных молекул ДНК, которые при внедрении в генетический аппарат придавали бы организму свойства, полезные для человека.
Технология рекомбинантных ДНК использует следующие методы:
- специфическое расщепление ДНК рестрицирующими нуклеазами, ускоряющее выделение и манипуляции с отдельными генами;
- быстрое секвенирование всех нуклеотидов очищенном фрагменте ДНК, что позволяет определить границы гена и аминокислотную последовательность, кодируемую им;
- конструирование рекомбинантной ДНК;
- гибридизация нуклеиновых кислот, позволяющая выявлять специфические последовательности РНК или ДНК с большей точностью и чувствительностью, основанную на их способности связывать комплементарные последовательности нуклеиновых кислот;
- клонирование ДНК: амплификация in vitro с помощью цепной полимеразной реакции или введение фрагмента ДНК в бактериальную клетку, которая после такой трансформации воспроизводит этот фрагмент в миллионах копий;
- введение рекомбинантной ДНК в клетки или организмы
Генетическая инженерия - потомок молекулярной генетики, но своим рождением обязана успехам генетической энзимологии и химии нуклеиновых кислот, так как инструментами молекулярного манипулирования являются ферменты. Если с клетками и клеточными органеллами мы подчас можем работать микроманипуляторами, то никакие, даже самые мелкие микрохирургические инструменты не помогут при работе с макромолекулами ДНК и РНК. Что же делать? В роли "скальпеля", "ножниц" и "ниток для сшивания" выступают ферменты.
Только они могут найти определенные последовательности нуклеотидов, "разрезать" там молекулу или, наоборот, "заштопать" дырку в цепи ДНК. Эти ферменты издавна работают в клетке, выполняя работы по репликации (удвоению) ДНК при делении клетки, репарации повреждений (восстановлению целостности молекулы), в процессах считывания и переноса генетической информации из клетки в клетку или в пределах клетки. Задача генного инженера - подобрать фермент, который выполнил бы поставленные задачи, то есть смог бы работать с определенным участком нуклеиновой кислоты.
Следует отметить, что ферменты, применяемые в генной инженерии, лишены видовой специфичности, поэтому экспериментатор может сочетать в единое целое фрагменты ДНК любого происхождения в избранной им последовательности. Это позволяет генной инженерии преодолевать установленные природой видовые барьеры и осуществлять межвидовое скрещивание.
Ферменты, применяемые при конструировании рекомбинантных ДНК, можно разделить на несколько групп:
- ферменты, с помощью которых получают фрагменты ДНК (рестриктазы);
- ферменты, синтезирующие ДНК на матрице ДНК (полимеразы) или РНК (обратные транскриптазы);
- ферменты, соединяющие фрагменты ДНК (лигазы);
- ферменты, позволяющие осуществить изменение структуры концов фрагментов ДНК.
Методы клонирования ДНК.Существует два подхода к клонированию ДНК. Первый подход предполагает использование бактериальных или дрожжевых клеток для размножения введенной в них чужеродной ДНК. Второй способ представляет собой амплификацию ДНК in vitro.
Клонирование ДНК in vivo
Используя микроорганизмы, можно создавать два типа библиотек ДНК: геномную и клоновую (кДНК).
Геномная библиотека. Если геном какого-либо организма разрезать, вставить в плазмидные или вирусные вектора и ввести в клетку, то в таком виде его можно сохранить. При разрезании плазмидной или фаговой ДНК вероятность выпадения целых и неизмененных кусков генома довольно высока.
Такой способ получения геномной библиотеки получил название «метод дробовика», так как геном в данном случае представлен отдельными фрагментами.
Принципы создания плазмидных и вирусных векторов общие, поэтому рассмотрим их на примере плазмидных. Следует отметить, что из вирусных ДНК лучше использовать ДНК фагов, так как они имеют большую емкость и позволяют вставлять более крупные куски генома.
Очищенные кольцевые молекулы ДНК обрабатывают рестриктазой, получая линейную ДНК. Клеточную ДНК обрабатывают той же рестриктазой, добавляют к плазмидной, добавляют лигазы. Таким образом получают рекомбинантную плазмидную ДНК, которую вводят в бактериальные или дрожжевые клетки. Плазмида реплицируется с образованием многих копий. Многие плазмиды несут ген устойчивости к антибиотикам, и если в рекомбинантной плазмиде есть такой ген, то клетки легко выявлять, выращивая на среде с антибиотиком.
Каждая такая колония представляет собой клон или потомство одной клетки. Плазмиды одной колонии содержат клон геномной ДНК, а совокупность плазмид можно назвать библиотекой геномной ДНК. Недостаток такого метода в том, что фрагменты ДНК образуются в огромном количестве. Разрезание геномной ДНК определяется случаем, поэтому лишь часть фрагментов содержат полноценные гены. Некоторые фрагменты могут содержать только часть гена или же интронные последовательности.
Библиотека кДНК. Создание кДНК начинается с синтеза на матрице РНК с помощью обратной транскриптазы комплементарной нити ДНК. Затем создают щелочные условия, разрушают цепь РНК на нуклеотиды, после чего с помощью ДНК-полимеразы синтезируют комплементарную цепь ДНК. При этом образуется фрагмент ДНК с тупыми концами. Такую ДНК встраивают в плазмиды и вводят в клетки бактерий. При амплификации плазмиды образуется клон комплементарной копии ДНК (кДНК).
Преимущества клоновой ДНК перед клонами геномной ДНК в том, что кодирующая белок нуклеотидная последовательность гена ничем не прерывается.
Гены эукариот содержат интроны, которые должны удаляться из транскриптной РНК перед превращением ее в матричную, после чего следует сплайсинг (сращивание). Бактериальные клетки не могут осуществлять такую модификацию РНК, образовавшуюся путем транскрипции гена эукариотической клетки. Поэтому если преследуют получение белка путем экспрессии клонированного гена, то лучше использовать банк кДНК, полученной на основе матричной РНК.
Полимеразная цепная реакция
В 1985 году К. Мюллис с сотрудниками разработали метод клонирования последовательностей ДНК in vitro, который получил название полимеразной цепной реакции (ПЦР).
К анализируемому образцу ДНК добавляют в избытке 2 синтетических олигонуклеотида - праймера размером около 20 нуклеотидов. Каждый из них комплементарен одному из 3’-концов фрагмента ДНК. ДНК нагревают для разделения цепей двойной спирали, а при охлаждении происходит гибридизация праймеров с комплементарными участками фрагментов ДНК. В результате в растворе будут находиться однонитевые ДНК с короткими двухцепочечными участками - затравками (праймерами). При добавлении нуклеотидов и ДНК-полимеразы синтезируются комплементарные цепи и образуются идентичные фрагменты ДНК (первый цикл, рис. 8). Реакция останавливается и ДНК снова денатурируется прогреванием.
В процессе охлаждения праймеры, находящиеся в избытке, вновь эффективно гибридизуются, но уже не только с цепями исходной ДНК, но и с вновь синтезированными. Внесение в систему ДНК-полимеразы инициирует второй цикл полимеразной реакции. Многократное повторение описанной процедуры позволяет провести 30 и более циклов ферментативного удлинения праймеров. При этом число сегментов ДНК, ограниченных с обоих концов используемыми праймерами, с каждым циклом ПЦР увеличивается экспоненциально (приближается к зависимости 2n, где n — число циклов). Выход всех других продуктов реакции увеличивается по линейной зависимости (рис. 9). Таким образом, в процессе рассматриваемой реакции эффективно амплифицируется только та последовательность ДНК, которая ограничена праймерами.

Рисунок 5. - Первый цикл полимеразной цепной реакции
Первоначально для ПЦР использовали фрагмент Кленова ДНК-полимеразы I E. coli. Однако недостатком данного подхода являлось то, что после каждого цикла реакции необходимо было вносить в реакционную смесь новую порцию фермента. Кроме того, в оптимальных температурных условиях такой полимеразной реакции (37 °С) появлялись вторичные участки связывания праймеров и наблюдалась амплификация незапланированных сегментов генома, т. е. специфичность амплификации не была полной. Существенное улучшение метода полимеразной цепной реакции было достигнуто после замены фрагмента Кленова на ДНК-полимеразу термофильной бактерии Thermus aquaticus (Taq-полимераза). Температурный оптимум реакции, направляемой Taq-полимеразой, находится в районе 70 °С. Другим важным свойством является то, что данная полимераза не инактивируется после длительной инкубации при 95 °С.

Рисунок 6. - Схема полимеразной цепной реакции
Используя Taq-полимеразу, удалось решить сразу две проблемы. Во-первых, термостабильная полимераза не инактивируется на этапе денатурации ДНК, и поэтому нет необходимости после каждого цикла реакции добавлять новую порцию фермента. Такое упрощение процедуры позволило автоматизировать проведение ПЦР, так как теперь требовалось лишь перенесение образца с определенным интервалом времени в разные температурные условия: 90—95 °С (температура денатурации) и 60—70 °С (температура ренатурации ДНК и ферментативной реакции). Во-вторых, высокий температурный оптимум реакции, катализируемой Taq-полимеразой, позволяет подбирать жесткие температурные условия отжига, обеспечивающие гибридизацию праймеров только в заданном районе изучаемого генома, что существенно повышает специфичность и чувствительность метода.
Используя метод ПЦР, можно in vitro селективно обогащать препарат ДНК фрагментом с определенной последовательностью в миллион и более раз. Это позволяет надежно выявлять однокопийные гены и их варианты в таких больших и сложных геномах, каким является геном человека. Чувствительность метода такова, что амплифицировать в ПЦР и выявить целевую последовательность можно даже в том случае, если она встречается однажды в образце из 105 клеток. Получаемый сегмент ДНК надежно выявляется в виде дискретной полосы после электрофоретического разделения молекул ДНК и окраски их этидиум бромидом. Если к праймеру пришить фермент, то ферментная метка будет накапливаться при амплифицировании. Продукт амплификации проверяется по принципу ИФА, то есть добавляется субстрат и отмечается изменение окраски. В качестве метки можно использовать стрептавидин. Его можно пришивать как к праймеру, так и к нуклеотидам. В последнем случае нуклеотиды, меченные стрептавидином, добавляются к обычным, идущим на синтез комплементарной цепи ДНК. Этим достигается еще большее усиление сигнала.
Размноженный in vitro фрагмент получают в количествах, достаточных для его прямого секвенирования. Поскольку при этом не требуется промежуточный этап клонирования фрагмента ДНК в молекулярных векторах, ПЦР иногда называют бесклеточным молекулярным клонированием (cell-free molecular cloning). Автоматизированная процедура Taq-полимеразной цепной реакции, состоящая из 30 и более циклов, занимает 3—4 часа, что существенно быстрее и проще процедуры клонирования определенного фрагмента ДНК в составе векторных молекул.
Применение метода полимеразной цепной реакции
Используя метод ПЦР, можно in vitro селективно обогащать препарат ДНК фрагментом с определенной последовательностью в миллион и более раз. Это позволяет надежно выявлять однокопийные гены и их варианты в таких больших и сложных геномах, каким является геном человека.
Чувствительность метода такова, что амплифицировать в ПЦР и выявить целевую последовательность можно даже в том случае, если она встречается однажды в образце из 105 клеток. Получаемый сегмент ДНК надежно выявляется в виде дискретной полосы после электрофоретического разделения молекул ДНК и окраски их этидиум бромидом. Если к праймеру пришить фермент, то ферментная метка будет накапливаться при амплифицировании. Продукт амплификации проверяется по принципу ИФА, то есть добавляется субстрат и отмечается изменение окраски. В качестве метки можно использовать стрептавидин. Его можно пришивать как к праймеру, так и к нуклеотидам. В последнем случае нуклеотиды, меченные стрептавидином, добавляются к обычным, идущим на синтез комплементарной цепи ДНК. Этим достигается еще большее усиление сигнала.
Размноженный in vitro фрагмент получают в количествах, достаточных для его прямого секвенирования. Поскольку при этом не требуется промежуточный этап клонирования фрагмента ДНК в молекулярных векторах, ПЦР иногда называют бесклеточным молекулярным клонированием (cell-free molecular cloning). Автоматизированная процедура Taq-полимеразной цепной реакции, состоящая из 30 и более циклов, занимает 3—4 часа, что существенно быстрее и проще процедуры клонирования определенного фрагмента ДНК в составе векторных молекул.
Полимеразную цепную реакцию используют для анализа индивидуальных вариаций последовательности нуклеотидов определенных локусов, для повышения эффективности клонирования целевых последовательностей изучаемых геномов и их прямого секвенирования, для детекции в организме патогенных микроорганизмов и т. п. (рис.10).
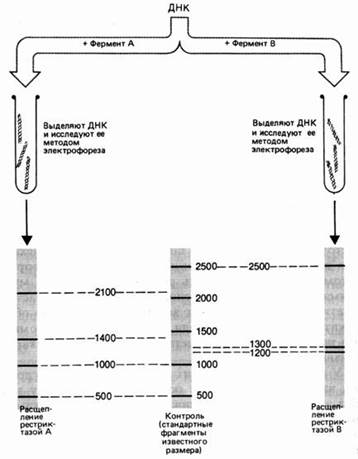
Рисунок 7. - Результаты электрофореза после обработки фрагмента ДНК разными рестриктазами
Используя 32Р-меченные синтетические олигонуклеотиды, можно выявлять единичные замены нуклеотидов в выбранных локусах геномной ДНК человека (или других организмов). Для этого в обычном варианте метода исследуемую ДНК гидролизуют рестриктазами, фракционируют электрофорезом, переносят разделенные фрагменты по Саузерну на нитроцеллюлозный фильтр, который гибридизуют с данным меченым олигонуклеотидом в условиях, при которых даже точечная замена нуклеотидов в анализируемой последовательности приводит к разрушению комплекса ДНК-олигонуклеотид.
Использование полимеразной цепной реакции для амплификации анализируемого локуса позволяет существенно упростить рассмотренный подход и повысить его чувствительность и специфичность. При этом для анализа аллельных вариантов генов достаточно всего 1 нг геномной ДНК человека, а гибридизацию можно проводить с негидролизованной рестриктазами ДНК, иммобилизованной на нитроцеллюлозном фильтре в виде небольшого пятна. Такой вариант метода позволил разработать новые диагностические тесты на генетические и инфекционные заболевания. В частности, этот подход используют для ранней диагностики наличия в организме вируса иммунодефицита человека (ВИЧ), что не удается осуществить другими методами. При этом не требуется работать с радиоактивными изотопами, так как амплифицированный сегмент вирусной ДНК выявляется напрямую после электрофоретического разделения ДНК и окраски их бромистым этидием.
Метод ПЦР позволил проанализировать наличие последовательностей вирусов папилломы человека в срезах биопсий новообразований шейки матки человека, залитых парафином за 40 лет до данного исследования. Более того, с помощью ПЦР удалось амплифицировать и клонировать фрагменты митохондриальной ДНК из ископаемых останков мозга человека возраста 7 тысяч лет!
На лизатах индивидуальных сперматозоидов человека продемонстрирована возможность одновременно анализировать два локуса, расположенных на разных негомологичных хромосомах. Такой подход обеспечивает уникальную возможность тонкого генетического анализа и изучения хромосомной рекомбинации, ДНК-полиморфизма и др. Метод анализа индивидуальных сперматозоидов сразу нашел практическое применение в судебной медицине, так как HLA-типирование гаплоидных клеток позволяет определять отцовство или выявлять преступника (комплекс HLA представляет собой набор генов главного комплекса гистосовместимости человека; локусы комплекса HLA — наиболее полиморфные из всех известных у высших позвоночных: в пределах вида в каждом локусе существует необычайно большое число разных аллелей — альтернативных форм одного и того же гена).
Используя ПЦР, можно выявлять правильность интеграции чужеродных генетических структур в заранее определенный район генома изучаемых клеток. Суммарная клеточная ДНК отжигается с двумя олигонуклеотидными затравками, одна из которых комплементарна участку хозяйской ДНК вблизи точки встраивания, а другая — последовательности интегрированного фрагмента в антипараллельной цепи ДНК. Полимеразная цепная реакция в случае неизмененной структуры хромосомной ДНК в предполагаемом месте встройки приводит к образованию фрагментов одноцепочечной ДНК неопределенного размера, а в случае запланированной встройки — двухцепочечных фрагментов ДНК известного размера, определяемого расстоянием между местами отжига двух праймеров. Причем степень амплификации анализируемого района генома в первом случае будет находиться в линейной зависимости от количества циклов, а во втором — в экспоненциальной. Экспоненциальное накопление в процессе ПЦР амплифицируемого фрагмента заранее известного размера позволяет визуально наблюдать его после электрофоретического фракционирования препарата ДНК и делать однозначное заключение о встройке чужеродной последовательности в заданный район хромосомной ДНК.
Введение нового гена в клетку
Ввести рекомбинантный ген в клетку можно 2 способами: используя вектора или путем прямого введения.
Требования к векторной ДНК, ее состав
Вектор - молекула ДНК или РНК, состоящая из двух компонентов: векторной части (носителя) и клонируемого чужеродного гена. Задача вектора – донести выбранную ДНК в клетку-рецепиент, встроить ее в геном, позволить идентификацию трансформированных клеток, обеспечить стабильную экспрессию введенного гена.
Таким образом, вектор должен быть небольшим, способным поддерживаться в клетке-хозяине (реплицироваться), многократно копироваться (ампфлицироваться), экспрессировать соответствующий ген (содержать соответствующие регуляторные последовательности), должен иметь маркерный ген, позволяющий различать гибридные клетки для эффективной селекции их; должен быть способен передаваться в клетку соответствующего организма.
Регуляторные последовательности, отвечающие за стабильную экспрессию гена, будут рассмотрены позднее.
Можно выделить 2 группы маркерных генов, позволяющие отличить трансформированные клетки:
1.Селективные гены, отвечающие за устойчивость к антибиотикам (канамицину, тетрациклину, неомицину и др.), гербицидам (у растений). Это могут быть гены ауксотрофности по какому-либо субстрату и т.д. Основной принцип работы такого маркера – способность трансформированных клеток расти на селективной питательной среде, с добавкой определенных веществ, ингибирующих рост и деление нетрансформированных, нормальных клеток.
2. Репортерные гены, кодирующие нейтральные для клеток белки, наличие которых в тканях может быть легко тестировано.
Чаще всего в качестве репортерных используются гены β-глюкуронидазы (GUS), зеленого флюоресцентного белка (GFP), люциферазы (LUC), хлорамфениколацетилтрансферазы (CAT). К настоящему времени из этого арсенала наиболее часто используют гены GUS и GFP и, в меньшей степени, LUC и CAT. Используемый в настоящее время как репортерный ген GUS является модифицированным геном из Escherichia coli, кодирующим β-глюкуронидазу с молекулярной массой 68 кД. GUS активен в широком диапазоне условий среды с оптимумом при рН 5-8 и 37°С. Он может гидролизовать обширный спектр природных и синтетических глюкуронидов, что позволяет подбирать соответствующие субстраты для спектрофотометрического или флюориметрического определения активности фермента, а также для гистохимического окрашивания тканей in situ (например, в синий цвет). Фермент достаточно стабилен: он устойчив к нагреванию (время полужизни при 55°С составляет около 2 ч) и к действию детергентов. В процессе замораживания-оттаивания потери активности GUS не происходит. В составе химерных белков, созданных генно-инженерными методами, GUS обычно сохраняет свою функциональную активность. В живых клетках белок GUS также весьма стабилен и активен от нескольких часов до нескольких суток.
GFP (green fluorescent protein - зеленый флюоресцентный белок, или белок зеленой флюоресценции) был обнаружен Shimomura с соавт. в 1962 г. у люминесцирующей медузы Aequorea victoria. Ген GFP был клонирован в 1992 г. Prasher и соавт., и уже через несколько лет началось активное использование этого гена как репортерного в работах с самыми разными про- и эукариотическими организмами. В настоящее время ген GFP применяется в сотнях работ во всем мире, и число их стремительно нарастает. Столь быстрый рост вызван особыми свойствами белка GFP, а именно его способностью флюоресцировать в видимой (зеленой) области спектра при облучении длинноволновым УФ. Эта флюоресценция обусловлена непосредственно белком, для ее проявления не требуется субстратов или кофакторов. Благодаря этому свойству ген GFP является очень перспективным репортерным геном, позволяющим проводить разнообразные прижизненные (недеструктивные) исследования с трансгенными организмами.
Многочисленные производные GFP получили общее название AFP (autofluorescent proteins - автофлюоресцентные белки). Из морской анемоны Discosoma sp. недавно выделен еще один белок DsRed, флуоресцирующий в красном свете. Еще несколько аналогичных флюоресцирующих белков было выделено в самое последнее время учеными Российской академии наук из различных коралловых полипов порядка Anthozoa. Он может быть денатурирован очень высокой температурой, крайними значениями рН или сильными восстановителями типа Na2SO4. При возвращении к физиологическим условиям GFP в значительной степени восстанавливает способность к флюоресценции. В составе химерных белков, созданных генноинженерными методами, GFP обычно сохраняет свою функциональную активность. В живых клетках белок GFP также очень стабилен.
CAT – гены отвечают за синтез хлорамфениколацетилтрансферазы (выделены из Escherihia coli). Этот фермент катализирует реакцию переноса ацетильной группы от ацетил-КоА к хлорамфениколу. Определяется гистохимически, по изменению окраски ткани при добавлении соответствующего субстрата.
LUC – ген кодирует фермент люциферазу (клонирована из бактерий и светлячка). Она вызывает свечение трансформированных клеток. Бактериальный фермент состоит из двух субъединиц. Для определения активности ферментов необходимо специальное оборудование - флуориметр и цифровая видеокамера с амплификатором светового сигнала. Фермент теряет активность при действии детергентов и повышенной температуры. Замена селективных генов на репортерные при отборе трансгенных растений часто весьма желательна, так как возможность потенциального риска для окружающей среды и здоровья человека при использовании репортерных генов практически исключена. Однако область применения репортерных генов шире, чем просто контроль трансгеноза. Другое, и, очевидно, более важное назначение репортерных генов состоит в том, чтобы выявлять (по возможности количественно) временные и пространственные особенности экспрессии данного конкретного гена, будь то собственного или чужеродного. Присоединение репортерного гена к одной лишь промоторной области позволяет исследовать в "чистом виде" ее роль в регуляции экспрессии изучаемого гена на уровне транскрипции.
Замена белок-кодирующей области гена на репортерную при сохранении участка, кодирующего 5'-концевую не транслируемую последовательность мРНК, позволяет оценить роль этой последовательности в процессах транспорта мРНК из ядра в цитоплазму и инициации трансляции
Одно из самых важных свойств гена - способность к экспрессии. За это свойство отвечают различные генетические элементы, которые мы должны встроить в векторную молекулу, несущую ген.
Типы векторов для введения гена в клетку
Существует несколько типов векторов:
1Бактериальные плазмиды Основная масса клеточной ДНК бактерий содержится в хромосоме (в хромосоме E. coli, например, 4 млн. пар нуклеотидов). Однако кроме хромосом бактерии содержат большое количество очень маленьких кольцевых молекул ДНК плазмид длиной несколько тысяч пар оснований (молекулярная масса от 1,5 до 300 мегадальтон, 1 МД = 1500 п.о). Такие мини-хромосомы называют плазмидами. Как правило, плазмиды имеют в своем составе гены устойчивости к антибиотикам, ионам тяжелых металлов (R-плазмиды), а также гены, контролирующие катаболизм некоторых органических соединений (плазмиды биодеградации, или D-плазмиды). Поскольку эти гены находятся в плазмидах, они представлены гораздо большим числом копий. Высокая копийность плазмид обеспечивает клетке синтез большого количества ферментов, химически нейтрализующих антибиотики или ксенобиотики, что и обеспечивает устойчивость к последним. Плазмиды, по-видимому, вездесущи, так как их выделяют из разных штаммов и видов бактерий, но не являются обязательными компонентами генома, а в некоторых природных штаммах плазмиды не обнаружены вообще.
Поскольку плазмидная ДНК значительно меньше хромосомной, ее довольно легко выделить в чистом виде. В присутствии ионов кальция плазмиды легко поглощаются бактериями-рецепиентами, даже если те их никогда не содержали, и в клетках бактериального потомства можно обнаружить много копий поглощенной плазмиды. Однако бактериальная клетка обычно может содержать в своем составе плазмиды одного типа. Это явление несовместимости плазмид. Существуют группы несовместимости – Inc-группы (от английского incompatibility – несовместимость). В такой группе может быть несколько плазмид, совместимых между собой, но не совместимых с другими плазмидами. У этих плазмид сходны многие признаки и часто значительна гомология ДНК.
Число копий плазмиды в клетке может существенно варьировать. Это зависит от генетических особенностей как клетки, так и плазмиды. Плазмиды, находящиеся "под ослабленным контролем", могут размножаться до тех пор, пока их количество не достигнет 10-200 копий на клетку. Если же плазмида находится "под строгим контролем", она реплицируется с той же скоростью, что и главная хромосома. Такие плазмиды содержатся в клетке в одной или в нескольких копиях. Естественно, что для клонирования рекомбинантных ДНК стараются использовать плазмиды первого типа. Но это не обязательно, так как плазмиды в присутствии хлорамфеникола могут умножаться независимо от деления хромосомы, и количество копий плазмиды может многократно увеличиваться.
Одна их наиболее часто употребляемых плазмид для клонирования pBR 322 создана на основе плазмид природного происхождения, выделенных из E. coli. Эта плазмида содержит гены устойчивости к двум антибиотикам: ампициллину и тетрациклину, причем в генах устойчивости к этим антибиотикам имеются сайты рестрикции. Если фрагмент чужеродной ДНК встраивается в один из генов устойчивости, то последний инактивируется. Следовательно, успешное встраивание фрагмента чужеродной ДНК в один из этих генов легко детектировать по исчезновению у бактерий устойчивости к данному антибиотику. Но при этом сохраняется устойчивость к другому антибиотику. Таким образом вектор дает возможность детектировать только те клоны бактерий, которые содержат рекомбинантную плазмиду.
2.Вирусы Есть вирусы, которые не ведут к гибели клетки, но встраиваются в геном клетки-хозяина и размножаются вместе с ней, либо вызывают ее неконтролируемый рост, т.е. превращают в раковую. К таким относятся ДНК-вирусы SV-40 и вирус полиомы. Внедрение некоторых опухолевых РНК-вирусов ведет к отпочковыванию вирусных частиц от клетки без ее лизиса. К таким вирусам относятся, например, ретровирусы (вирус саркомы Рауса и СПИДа). Для бактериальных клеток в качестве вектора часто используют бактериофаги.
Вирусы являются одними из главных кандидатов на роль векторов для введения чужеродной ДНК. При вирусной инфекции каждая клетка может получить большое число копий чужеродного гена. ДНК можно встраивать так, чтобы она находилась под контролем сильных вирусных промоторов, что обеспечит высокий уровень экспрессии гена, и его продукты будут более доступны для исследования.
В последние годы сконструированы многочисленные "челночные" векторы и их рекомбинантные производные, способные к репликации в животной и бактериальной клетке и эффективно экспрессирующие клонируемый ген в животной клетке. Наиболее распространенные векторы состоят из плазмиды рВR322 и интактного раннего района транскрипции ДНК SV40, а нужный ген встраивается под контроль промотора поздних генов или дополнительного раннего промотора. Например, в ДНК SV40 был встроен ген β-глобина кролика, который экспрессировался в линии клеток обезьяны, зараженных рекомбинантным вирусом: в клетках синтезировались и мРНК гена глобина, и сам белок.
Вирус должен быть жизнеспособным после рекомбинирования его ДНК. Легче всего вирусы вводятся в бактерии. Недостатком вирусов как векторов является их небольшая емкость. Кроме того, вирусы заражают небольшой круг хозяев.
3.Существуют гибридные вектора, содержащие ДНК фага и плазмиды. К ним относятся, например, космиды и фазмиды.
Космиды – плазмидные вектора, в которые встроен участок генома фага λ, обеспечивающий возможность упаковки этой молекулы ДНК в фаговую частицу. Фаговые частицы обеспечивают хорошее проникновение гибридной ДНК в клетку (путем инъекции), после чего происходит замыкание ДНК в кольцо по липким концам и репликация ее по плазмидному типу.
Фазмиды также являются гибридами между фагом и плазмидой. После встройки чужеродной ДНК могут в одних условиях развиваться как фаги, в других – как плазмиды.
4. Вироиды
Из всех известных в настоящее время инфекционных агентов имеют ранг наиболее странных. Известно, что самые мелкие вирусы, способные к независимой репликации, имеют размеры генома, соответствующие молекулярной массе 1 М, то есть около 1500 тыс. пар оснований. Это считали минимальным количеством генетической информации, необходимой для кодирования вирусоспецифических продуктов и подавления метаболизма хозяйской клетки.
Однако в 1971 году были открыты инфекционные агенты, представляют собой очень короткую цепь 1 нитевой ковалентно связанной кольцевой РНК, состоящую из 270-300 нуклеотидов (на три порядка меньше самых минимальных вирусов), не заключенную в белковую оболочку. Это необычные патогены - самые простые и самые маленькие из всех известных.
Каким образом вироиды продуцируют симптомы болезни в инфицированных растениях, не известно до сих пор. Установлено, что они реплицируются ферментами клетки-хозяина, не транслируются в видоспецифичные полипептиды, интегрируются в геном клетки-хозяина.
Вироиды заражают персиситентно (не происходит выздоровления). Вызывают системную инфекцию, т.е. мигрируют из сайта внедрения в другие части растений, переносятся механически или через клеточный сок, через семена, пыльцу. Вироиды также связаны с ядерными фракциями растений и могут размножаться в ядрах.
При работе с вироидами получают 1-нитевую ДНК- копию РНК и достраивают комплементарную нить для получения 2-нитевой ДНК вироида. Такая 2-цепочечная ДНК вcтраивается в плазмиду и передается в клетки E. coli для клонирования. Считывание гена начинается с промотора, который узнается РНК-полимеразой, отвечающей за транскрипцию ДНК в матрицу РНК. Обычно это фрагмент ДНК из 41-44 пар оснований. Ген считывается слева направо, от 5’ к 3’ концу гена и заканчивается в терминальной области гена. За промотором начинается стартовый сайт транскрипции, за которым следует смысловая часть гена. Промоторная область гена содержит определенные короткие сочетания нуклеотидов, характерные для бактериальных генов, или для генов высших организмов. Такие сочетания служат сигналами для РНК-полимеразы, которая присоединяется к промоторной части гена и начинает его считывать.
Однонитевые и двунитевые ДНК способны инициировать репликацию вироида в механически инокулированных растениях табака. Энзиматически in vitro синтезированы также РНК вироидов, высокоинфекционные для растений. Векторные системы могут быть разработаны на основе самих РНК, на основе вироидоспецифичных ДНК, а также в комбинации вироидоспецифичных ДНК с Ti-плазмидами. Вироиды инфицируют своих хозяев в течение всего их жизненного цикла, поэтому в случае использования вироидных векторных систем можно ожидать постоянной экспрессии чужеродного гена в растении.
5.Плазмиды агробактерий
В качестве векторов могут использоваться опухолеобразующие плазмиды бактерий. Виды Agrobacterium эволюционно родственны клубеньковым бактериям, относящимся к роду Rhizobium, и имеют много общих с ними черт. Однако характер взаимодействия агробактерий с растением имеет своеобразные особенности.
Взаимодействие видов Agrobacterium с растениями представляет особый интерес, так как при этом виде паразитизма один из партнеров специфически видоизменяет свойства хозяина, встраивая свои гены в его геном. Кроме того, это служит уникальным примером миграции ДНК прокариот в эукариотическую клетку. ДНК митохондрий и хлоропластов Хлоропласты и митохондрии содержат полноценную генетическую систему, то есть все компоненты, необходимые для экспрессии генетической информации: ДНК, ДНК-полимеразы, РНК-полимеразы и белоксинтезирующий аппарат (рибосомы, т-РНК, аминоацил-тРНК-синтетазы).
6.Хлоропластная и митохондриальная ДНК также привлекают внимание ученых в качестве возможных векторов для переноса генов в клетку. Структурная организация этих клеточных субгеномов существенно различается.
Хлоропласты и другие пластиды обладают одинаковой генетической информацией, так называемым пластомом. У высших растений он представляет собой замкнутую молекулу ДНК длиной 150 т. н. п., достаточную для кодирования примерно 100 белков. Для синтеза пластид необходимо значительно больше белков. Остальные белки кодируются ядром, синтезируются в цитоплазме и поступают в хлоропласты. Некоторые важнейшие белки хлоропластов состоят из нескольких субъединиц, часть из них синтезируется на рибосомах цитоплазмы и транспортируется в хлоропласт, где они объединяются с другими полипептидами, закодированными в самом хлоропласте и там же синтезируемыми. Таким образом, для биосинтеза функционально активного хлоропласта требуется согласованная экспрессия генома и пластома.
Различные типы пластид содержат неодинаковые количества идентичных копий пластома: от 10 – 20 копий в пластидах корней и зрелых хлоропластах до сотен копий в молодых хлоропластах картофеля. Такой уровень амплификации позволяет надеяться на надежную экспрессию чужеродной ДНК при использовании их в качестве векторов в генноинженерных экспериментах. Кроме того, гены рибосомальной РНК пластид и большой субъединицы РБФК кодируются геномом хлоропластов. Возможно, введение сильных промоторов в эти гены и дополнительная их модификация существенно повлияют на фотосинтетическую активность растительной ткани.
Гены растений также способны к экспрессии в клетках Е. coli. Это гены большой субъединцы РБФК. Преимущество хлоропластных генов заключается в том, что их экспрессия к клетках кишечной палочки может быть достигнута путем простого объединения транскрибируемых последовательностей, т.к. в ДНК хлоропластов и бактерий до начала стартовых кодонов трансляции расположена одинаковая нуклеотидная последовательность. Это дает возможность синтезировать растительные экономически важные полипептиды с помощью клеток прокариот.
В отличие от хлоропластной, ДНК митохондрий характеризуются исключительным разнообразием и их величина колеблется от 200 до 2400 т. н. п.. Однако никакой корреляции между размером митохондриального генома и числом белковых продуктов, синтезируемых изолированными митохондриями, не наблюдается. Это явление, а также большие размеры митохондриальной ДНК, по-видимому, можно объяснить присутствием ДНК, бесполезной для функционирования митохондрий.
В составе митохондриальной ДНК имеются структурные гены, кодирующие полипептиды, гены рибосомных и транспортных РНК. Однако большая часть белков митохондрий, как и хлоропластов, кодируется ядерными генами. Но если геном хлоропластов представлен гомогенной популяцией крупных кольцевых молекул, то в митохондриях содержится несколько классов кольцевых молекул, не все функции которых еще ясны.
Митохондриальный геном животных организмов намного меньше, 15 – 19 т. н. п., и более консервативен по структуре. Гены митохондрий кодируют 2 группы признаков – работу дыхательных систем и устойчивость к антибиотикам и другим ядам. В митохондриальном геноме растений есть также гены, отвечающие за признак мужской стерильности цитоплазмы.
7.Транспозоны
Транспозоны - сегменты ДНК, которые контролируют собственную транспозицию (перемещение) из одного сайта ДНК в другой путем вырезания из исходного сайта и внедрения в новый сайт хромосомы или плазмиды. Впервые были открыты в 40-х годах американской ученой Барбарой Мак-Клинток у кукурузы. Эти гены, индентифицированные по их способности подавлять экспрессию других генов кукурузы, находящихся рядом с ними, не имели фиксированного положения в хромосоме. Они как бы передвигались по всему геному растения. Регуляторные элементы могли встраиваться и выщепляться, причем после их выщепления зачастую начинали функционировать ранее молчащие гены.
Оказалось, что гены, ассоциированные с регуляторными элементами, становились нестабильными и часто мутировали из-за нестабильности самих этих элементов. В течение многих лет кукуруза оставалась единственной системой, в которой обнаруживались такие подвижные генетические элементы. Сейчас - и у бактерий, дрозофил и других организмов.
Механизм перемещения фрагментов ДНК по геному до конца не выяснен. ДНК переносится ферментом транспозазой. Фермент кодируется последовательность длиной около 20 нуклеотидов в середине транспозона. Он специфически взаимодействует с концевыми инвертированными повторами мобильного элемента и может вырезать его из хромосомы. Вырезание может происходить точно – с восстановлением исходной структуры участка ДНК, и неточно, то есть с делециями и вставками от одного до нескольких нуклеотидов. Это приводит к появлению стабильных мутаций и является одним из механизмов создания новых последовательностей ДНК.
Как правило, мобильные генетические элементы многократно повторены в геноме и образуют гетерогенные семейства, члены которых диспергированы по хромосомам. Большая часть членов каждого семейства являются дефектными копиями и не кодируют какой-либо функции, хотя сохраняют способность к перемещению. Поведение транспозонов можно расценить как паразитическое. Длина их от 2 до 10 тысяч нуклеотидных пар. У высших эукариот на долю транспозонов приходится примерно 10% ДНК клетки. Большинство их перемещается изредка, но, так как их в клетке довольно много, транспозиция оказывает значительное влияние на разнообразие видов.
Биологический смысл перемещения отдельных сегментов ДНК:
- прерывание соответствующего гена, что ведет к эволюции;
- регуляция деятельности генов, так как транспозоны могут нести сигналы для начала считывания генов. В новых областях усиливают или запрещают работу гена.
Транспозоны также участвуют в горизонтальном переносе генов.
У бактерий были обнаружены 2 класса подвижных генов, различающихся по длине и сложности организации.
1. Инсерционные последовательности, или 1S элементы, имеющие длину около тысячи пар нуклеотидов и содержащие только ген, отвечающий за их перемещение.
2. Транспозоны, длиной от 3 до 20 т. н. п., состоящие из ряда дополнительных генов, отвечающих за устойчивость бактерий к различным токсическим веществам.
Поскольку подвижные гены могут перемещаться в пределах генома с одного места на другое, то они могут быть весьма эффективными векторами для передачи рекомбинантной ДНК. Генетическая трансформация с помощью векторов на основе транспозонов была впервые осуществлена на дрозофиле. С помощью транспозируемого элемента Р дрозофиле был передан ген, обуславливающий коричневую окраску глаз. Перенос генов при помощи транспозонов имеет большие преимущества, так как он происходит с высокой частотой и не влечет значительных перестроек интегрируемой ДНК. Кроме того, этим методом можно переносить достаточно большие фрагменты ДНК
Способы прямого введения гена в клетку
Прямое введение гена в клетку осуществляют несколькими способами:
1.Трансфекция
2.Микроинъекция
3.Электропорация
4.Метод «мини-клеток»
5.Упаковка в липосомы
6.Электронная пушка
При трансфекции ДНК адсорбируется на кристаллах фосфата кальция. Образуются частицы кальциевого преципитата. Они поглощаются клеткой путем фагоцитоза.
Для повышения эффективности трансформации к специфической ДНК, содержащей ген, по которому будет производится селекция, добавляется неспецифическая ДНК-носитель. Обычно для этой цели берут ДНК из тимуса теленка или спермы лосося. Часть ДНК связывается с мембраной и не попадает в клетки. ДНК акцептируют от 15 до 90% клеток. Через несколько суток после введения небольшая доля клеток способны экспрессировать чужеродные гены, но затем уровень экспрессии падает и более или менее стабильную трансформацию претерпевает 10-3 - 10-5 клеток.
Для трансфекции используется и ДЭАЭ-декстран, полимер, адсорбирующий ДНК. Эффект вхождения в клетки и время экспрессии высоки, но частота стабильной трансформации ниже, чем при использовании преципитата кальция. Частоту трансфекции увеличивает глицериновый шок (4 минуты в 15% растворе глицерина в НEPES-буфере).
В клетки можно вводить любой ген, если заранее лигировать его с клонированным селективным маркером. Однако дальнейшие исследования показали, что лигирование вне клетки не обязательно. Клетки, поглощающие селективный ген, вместе с ним поглощают и другую ДНК, имеющуюся в кальциевом преципитате. Таким образом, пользуясь методом котрансформации, практически любой клонированный сегмент ДНК можно ввести в культивируемые клетки эукариот, если включить эту ДНК вместе с селективным маркером в состав смеси для образования кальциевого преципитата.
Для трансфекции можно использовать хромосомы или фрагменты хромосом. Клетки-доноры блокируются на стадии митоза. Митотические хромосомы высвобождаются под воздействием осмотического шока и гомогенизации. Их очищают путем дифференциального центрифугирования. Хромосомы осаждают на поверхности клеток хлористым кальцием, а через несколько часов обрабатывают реагентом, способным перфорировать мембраны (например, глицерином).
Для обработки клеток-рецепиентов используются грубо очищенные препараты хромосом, так как хромосомы при этом разрушаются меньше всего. Количество хромосом для обработки 1 клетки ограничено. Лучше использовать не более 20 хромосом на 1 клетку-рецепиент, так как при высоких концентрациях хромосом в суспензии они агглютинируют. Рецепиентная клетка содержит фрагменты донорных хромосом, которые могут встраиваться в геном, могут реплицироваться самостоятельно. Во введенных фрагментах часто наблюдаются делеции.
Не все клетки способны к трансформации геномной ДНК с высокой частотой. Человеческие фибробласты эффективно включают плазмидную ДНК и почти не включают геномную.
Микроинъекция ДНК в клетки млекопитающих стала возможной с появлением прибора для изготовления микропипеток диаметром 0.1-0.5 микрона и микроманипулятора (рис. 11). Так, плазмиды, содержащие фрагмент вируса герпеса с геном тимидинкиназы (ТК) и плазмиду рВR322, были инъецированы в ТК--клетки и было показано, что ТК-ген проник в ядра и нормально в них реплицировался. Метод введения ДНК с помощью микроинъекций был разработан в начале 70-х годов Андерсоном и Диакумакосом. В принципе, при наличии хорошего оборудования можно за 1 час инъецировать 500-1000 клеток, причем в лучших экспериментах в 50% клеток наблюдается стабильная интеграция и экспрессия инъецированных генов. Преимущество описываемого метода заключается также в том, что он позволяет вводить любую ДНК в любые клетки, и для сохранения в клетках введенного гена не требуется никакого селективного давления.
Электропорация основана на том, что импульсы высокого напряжения обратимо увеличивают проницаемость биомембран. В среду для электропорации добавляют клетки и фрагменты ДНК, которые необходимо ввести в клетки (рис. 12). Через среду пропускают высоковольтные импульсы (напряжение 200 - 350 В, длительность импульса 54 мс), приводящие к образованию пор (электропробой) в цитоплазматической мембране, время существования и размер которых достаточны, чтобы такие макромолекулы, как ДНК, могли из внешней среды войти в клетку в результате действия осмотических сил. При этом объем клетки увеличивается.
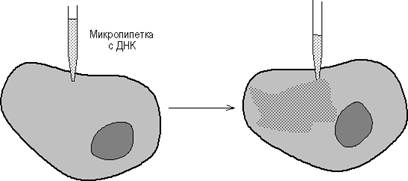
Рисунок 8. - Введение ДНК путем микроинъекции
Напряженность электрического поля и продолжительность его действия, концентрации трансформирующей ДНК и реципиентных клеток для каждой системы клеток подбирают экспериментально, с тем чтобы достичь высокого процента поглощения ДНК выжившими клетками. Показано, что в оптимальных условиях электропорации количество трансформантов может достигать 80% выживших клеток.
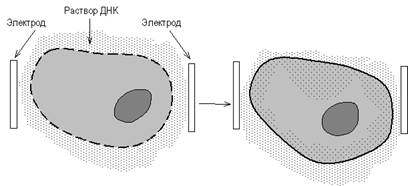
Рисунок 9. - Метод электропорации
Электропорация — физический, а не биохимический метод, и это, по-видимому, обусловливает его широкое применение. Многочисленные исследования продемонстрировали, что электропорация может успешно использоваться для введения молекул ДНК в разные типы клеток, такие как культивируемые клетки животных, простейшие, дрожжи, бактерии и протопласты растений. Электропорирующий эффект высоковольтного разряда на бислойную липидную мембрану, по-видимому, зависит от радиуса ее кривизны. Поэтому мелкие бактериальные клетки эффективно поглощают ДНК при значительно большей напряженности (10 кВ/см и более), чем крупные животные и растительные клетки, эффективно поглощающие ДНК при напряженности поля 1—2 кВ/см.
Электропорация — наиболее простой, эффективный и воспроизводимый метод введения молекул ДНК в клетки. Однако до недавнего времени этот метод использовался в ограниченном числе лабораторий в связи с отсутствием серийных приборов — электропораторов. Появление и совершенствование таких приборов в ближайшие годы приведет к широкому применению данного подхода в генетической инженерии самых разных типов клеток.
«Мини-клетки» получают путем блокирования донорных клеток митозе колцемидом. При продолжительной обработке клеток колцемидом в них вокруг каждой хромосомы формируется новая ядерная мембрана. Обработка цитохалазином В и центрифугирование приводит к образованию мини-клеток, представляющих микроядра, инкапсулированные в цитоплазматическую мембрану.
Полученные мини-клетки очень чувствительны к разного рода воздействиям, поэтому для слияния подбирают специальные мягкие условия. Метод трудный, капризный, эффективность низкая – 10-6 – 10-7.
Упаковка в липосомы используется для защиты экзогенного генетического материала от разрушающего действия рестриктаз.
Липосомы - сферические оболочки, состоящие из фосфолипидов. Получают их путем резкого встряхивания смеси водного раствора и липидов, либо обрабатывая ультразвуком водные эмульсии фосфолипидов. Липосомы, состоящие из фосфатидилсерина и холестерина наиболее пригодны для введения ДНК в клетки животных и растений. Системы переноса с помощью липосом низкотоксичны по отношению к клеткам.
Метод биологической баллистики (биолистики) является одним из самых эффективных на сегодняшний день методов трансформации растений, особенно однодольных.
Суть метода заключается в том, что на мельчайшие частички вольфрама, диаметром 0,6—1,2 мкм, напыляется ДНК вектора, содержащего необходимую для трансформирования генную конструкцию. Вольфрамовые частички, несущие ДНК, наносятся на целлофановую подложку и помещаются внутрь биолистической пушки. Каллус или суспензия клеток наносится в чашку Петри с агаризированной средой и помещается под биолистическую пушку на расстоянии 10—15 см. В пушке вакуумным насосом уменьшается давление до 0,1 атм. В момент сбрасывания давления вольфрамовые частички с огромной скоростью выбрасываются из биолистической пушки и, разрывая клеточные стенки, входят в цитоплазму и ядро клеток.
Обычно клетки, располагающиеся непосредственно по центру, погибают из-за огромного количества и давления вольфрамовых частиц, в то время как в зоне 0,6—1 см от центра находятся наиболее удачно протрансформированные клетки. Далее клетки осторожно переносят на среду для дальнейшего культивирования и регенерации.
С помощью биолистической пушки были протрансформированы однодольные растения, такие, как кукуруза, рис, пшеница, ячмень. При этом были получены стабильные растения-трансформанты. Кроме успехов в получении трансгенных однодольных, биолистическая трансформация применяется для прямого переноса ДНК в эмбриогенную пыльцу и дальнейшего быстрого получения трансгенных дигаплоидных растений, которые являются важным этапом в селекционной работе. В настоящее время этим методом была проведена трансформация растений табака и после регенерации гаплоидных растений получены стабильные трансформанты.
Генетические манипуляции с бактериальными клетками.В настоящее время бактерия E. coli является самой изученной клеткой из всех существующих. У большинства наиболее полно изученных фагов клеткой - хозяином является также E. coli.
Протопласт E. coli одет в муреиновый мешок, прилегающий к внешней мембране. E. coli относится к микроорганизмам, не обладающим физиологической компетентностью к поглощению экзогенной ДНК. Поэтому необходимо создать условия, позволяющие преодолеть барьер клеточной стенки. Сначала получают сферопласты путем обработки клеток лизоцимом в изотоническом растворе.
Липополисахаридный слой внешней мембраны грамотрицательной бактерии стабилизирован двухвалентными катионами, поэтому для разрыхления внешней мембраны E. coli используется комплексообразователь этилендиаминтетрауксусная кислота (EDTA), которая связывает двухвалентные катионы. При обработке EDTA часть липополисахаридов высвобождается из внешней мембраны клетки, и лизоцим может достигнуть муреинового мешка и гидролизовать его. Это ведет к повышению проницаемости клеточной оболочки. Усовершенствование методов получения сферопластов E. coli и их трансфекции позволили достичь достаточно высокой эффективности трансформации молекулами ДНК различных фагов.
Обнаружено, что на инфекционность существенное влияние оказывает форма молекул фаговых ДНК, которую они принимают in vivo. Фаги с кольцевой или линейной, но быстро замыкающейся ДНК (лямбоидные фаги) характеризуются наибольшей эффективностью трансфекции.
Успешное проведение экспериментов на кишечной палочке стало стимулом для проведения аналогичных исследований с другими прокариотическими организмами. Наибольших успехов удалось достичь с клетками Bacillus subtilis. B. subtilis - непатогенный почвенным микроорганизм, растущий в строго аэробных условиях. Бациллы не образуют токсинов и непатогенны ни для животных, ни для человека, тогда как клеточная стенка E. coli содержит эндотоксин, который довольно трудно отделить от продуктов генной инженерии. Кроме того, клеточная стенка бацилл имеет простую структуру и бактерии могут секретировать многие белки в культуральную жидкость. 20 различных видов бацил секретируют в культуральную жидкость более 40 ферментов с внеклеточной локализацией. E. coli секретирует в среду относительно мало белков, а выделение и очистка их затруднены. В бациллах также обнаружены плазмиды и фаги, которые к настоящему моменту уже хорошо изучены.
Чужеродные гены клонируют в так называемых челночных векторах. Эти вектора с одинаковым успехом реплицируются в клетках нескольких хозяев, в данном случае, в клетках E. coli и B.subtilis. Векторы были получены комбинацией in vitro фрагментов этих плазмид.
Гены E. coli со своими регуляторными районами не функционируют в B.subtilis, поэтому были использованы собственные гомологичные районы B.subtilis.
Для конструирования рекомбинантной ДНК, содержащей в своем составе ген, который должен экспрессироваться, придерживаются следующей стратегии. Синтезируют кДНК или из клонотеки выделяют клетки, несущие фрагмент генома с нужным геном, и клонируют их в соответствующем векторе. Фрагменты геномной ДНК подвергают модификации - удаляют из них некодирующие области и участки соседних генов. Часто для проведения этой операции необходимо секвенирование данного фрагмента ДНК. Затем конструируются промежуточные рекомбинантные ДНК, в которых ген помещается под контроль бактериальных регуляторных элементов (промотор, оператор, точка связывания с рибосомами). Эти регуляторные элементы выделяют из гибридных плазмид, сконструированных специально как источники регуляторных элементов. Полученная конструкция встраивается в подходящий вектор, например, pBR 322, и ген экспрессируется в бактериальной клетке.
Однако удобнее встраивать ген в специальный вектор для экспрессии, который уже содержит регуляторные элементы, обеспечивающие активную экспрессию после введения рекомбинантной плазмиды в бактериальную клетку. К таким эффективным регуляторным участкам относится, например, сильный промотор гена бэта-лактамазы (ген устойчивости к пенициллину, входящий в состав плазмиды pBR 322). Ряд генов, в том числе и ген инсулина, встраивали в сайт рестрикции Pst I, который расположен в структурной части гена. Промотор этого гена обеспечивает эффективную транскрипцию, которая продолжается до тех пор, пока РНК-полимераза не дойдет до сигнала терминации встроенного гена.
В качестве примера маркирования вектора могут служит первые эксперименты с E. coli, а точнее с одной из ее плазмид рBR322, проведенные Гилбертом для получения инсулина. Плазмида pBR322 содержит 2 гена, которые определяют устойчивость к ампициллину и тетрациклину. Рестриктаза PstI расщепляет плазмиду в средней части гена, кодирующего фермент устойчивости к апициллину. После расщепления плазмиды на ее концы с помощью концевой трансферазы надстраивали последовательность из четырех нуклеотидов с остатками гуанина. Затем, как обычно, с помощью лигаз "вшивали" ген проинсулина, получая рекомбинантную ДНК. Встроенный в плазмиду фрагмент ДНК нарушал синтез фермента, разрушающего ампициллин, но ген, обеспечивающий устойчивость к тетрациклину, оставался активным. Трансформированные таким образом клетки E. coli синтезировали гибридный белок, содержащий последовательности пенициллазы и проинсулина, поэтому биологически активный инсулин получали путем отщепления пенициллазы и средний сегмент проинсулина.
С другой стороны, если фрагмент чужеродной ДНК встраивается в один из генов устойчивости, то последний инактивируется. Следовательно, успешное встраивание фрагмента чужеродной ДНК в один из этих генов легко детектировать по исчезновению у бактерий устойчивости к данному антибиотику.
Введение генов в клетки млекопитающих.Манипуляции с клетками млекопитающих можно разделить на 2 большие группы: эксперименты с соматическим клетками и эксперименты по трансформации половых клеток. В последнем случае конечный результат – получение трансгенных организмом.
Характеристика векторов для переноса генов в животные клетки
Одними из лучших носителей для введения чужеродной информации в животную клетку являются вектора на основе ретровирусов, например, на основе вируса лейкоза мышей. Они обеспечивают высокоэффективный перенос генов и их стабильное встраивание в хромосому клеток-мишеней. В основном трансформации животных клеток осуществляют либо с помощью ретровирусов (около 40% от всех трансформаций), либо путем упаковки ДНК в липосомы (25%), реже используют аденовирусы, так как они могут вызывать сильный иммунный ответ, кроме того, невозможно их повторное введение.
Если проблема доставки чужеродной ДНК in vitro практически решена, а ее доставка в клетки-мишени разных тканей in vivo успешно решается (главным образом путем создания конструкций, несущих рецепторные белки, в том числе и антигены, специфичные для тех или иных тканей), то другие характеристики существующих векторных систем - стабильность интеграции, регулируемая экспрессия, безопасность — все еще нуждаются в серьезных доработках.
Прежде всего это касается стабильности интеграции. До настоящего времени интеграция в геном достигалась только при использовании ретровирусных либо аденоассоциированных векторов. Повысить эффективность стабильной интеграции можно путем совершенствования генных конструкций типа рецептор-опосредованных систем, либо путем создания достаточно стабильных эписомных векторов (то есть ДНК-структур, способных к длительной персистенции внутри ядер).
В последнее время особое внимание уделяется созданию векторов на базе искусственных хромосом млекопитающих (MAC - mammalian artificial chromosomes). Благодаря наличию основных структурных элементов обычных хромосом такие мини-хромосомы длительно удерживаются в клетках и способны нести полноразмерные (геномные) гены и их естественные регуляторные элементы, которые необходимы для правильной работы гена, в нужной ткани и в должное время. Такие искусственные хромосомы уже созданы для дрожжей (YAK), так как геном дрожжей полностью картирован.
Для идентификации модифицированных клеток, необходимы маркеры. Если трансформируют соматические клетки, то применяют обычно селективные маркеры. Аксель с коллегами из колледжа терапии и хирургии Колумбийского университета исправили таким образом генетический дефект клеток мыши. Они взяли фрагмент ДНК, содержащий ген тимидинкиназы (ТК), который получен из вируса герпеса, смешали эту ДНК с несколькими миллиграммами ДНК-носителя из спермы лосося и осадили ДНК на культуру L-клеток мыши, в которых ген ТК отсутствовал (ТК-). С частотой 1 на 100000 клетки приобретали ген ТК, поэтому на селективной среде, которая не позволяла расти ТК- - клеткам, росли и нормально размножались ТК+ - клетки.
Другой селективный маркер - ген, кодирующий дигидрофолатредуктазу (ДГФР), можно использовать при трансформации немутантных линий клетки. Благодаря экспрессии многих копий этого гена животная клетка вместе с плазмидой приобретает устойчивость к высоким концентрациям ингибитора фермента, и таким образом трансформантов можно отбирать при высоких концентрациях ингибитора.
Разработано еще два универсальных вектора, содержащих генные маркеры, работающие в нормальных клетках. Они построены по одному и то
Дата добавления: 2015-07-14; просмотров: 6488;