Введение ДНК в клетки растений с помощью Ti- и Ri-плазмид.
A. tumefaciens вызывает образование опухолей стебля двудольных растений - так называемых корончатых галлов. Бактерии прикрепляются к клетками растения в местах повреждений. Сайтами связывания на поверхности бактерий, видимо, являются молекулы β-глюкана и О-антигенной цепи липополисахарида внешней мембраны.
Бактерии связываются с рецепторами высшего растения, состоящими из белка и пектина; лектины в данном случае не имеют значения. Бактериальные сайты связывания и рецепторы растений являются констуитивными, т.е. оба партнера обладают ими еще до момента взаимодействия. Первый шаг взаимодействия с растением - узнавание - следует рассматривать как специфическую адгезию растений. Как только бактерии прикрепились к поверхности клеток растения, они начинают образовывать целлюлозные фибриллы. Эти фибриллы можно увидеть в сканирующем электронном микроскопе уже через 90 минут после добавления бактерий к суспензии культуры клеток ткани моркови. К 10 часам инкубации фибриллы формируют сеть, покрывающую поверхность растительных клеток. Фибриллы служат более прочному закреплению бактерий на поверхности хозяина. За целлюлозные фибриллы могут зацепиться свободно плавающие клетки бактерий. Фиксируя их у поверхности растения, фибриллы увеличивают множественность заражения. В результате размножения образуются скопления бактерий на поверхности растения.
Клеточная стенка растения повреждается вследствие выделения бактериями пектолитических ферментов, что обеспечивает плотный контакт бактерий с плазмалеммой растительной клетки. Этот контакт необходим для передачи ДНК от бактерий в растительную клетку. Передача ДНК происходит без нарушения целостности мембраны растительной клетки, но требует определенного её состояния - компетентности.
Способность A. tumefaciens индуцировать у растений образование опухолей типа "корончатого галла" коррелирует с наличием у них Ti-плазмиды. Опухолевая трансформация проявляется в гипертрофии возникающей после проникновения агробактерий в пораненные участки (сайты) растений (рис. 13). Трансформация является результатом стабильного ковалентного включения (инсерции или интеграции) сегмента («transferred» или Т-ДНК) большой плазмиды (pTi - tumor inducing или pRi — root inducing) бактерий в ядерную ДНК растительной клетки.
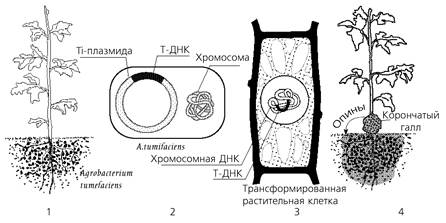
Рисунок 10. - Генетическая колонизация растения A. tumefaciens: 1- агробактерии существуют в ризосфере; 2 - строение A. tumefaciens; 3 – встраивание Т-ДНК в геном; 4 – образование опухоли
Другой вид агробактерий – A. rhizogenes, - вызывает заболевание, именуемое "бородатый корень", при котором в зоне повреждения корня образуется масса новых корешков. A. rubi обычно индуцируют неорганизованные опухоли (тератомы), штаммы A. radiobacter авирулентны.
В отличие от большинства тканей взятых из нормальных растений, трансформированные ткани в культуре in vitro в асептических (стерильных) условиях способны неограниченно расти в отсутствие экзогенно добавленных ауксинов и цитокининов. Кроме того, трансформированные ткани часто синтезируют одну или более групп соединений, названных опинами, которые обычно не обнаруживаются в нетрансформированных растительных тканях. Наиболее подробно изучены опухоли — корончатые галлы, индуцируемые Agrobacterium tumefaciens. Они представляют собой истинно злокачественные опухоли, которые могут расти в культуральной среде в отсутствие стимуляторов роста — фитогормонов, необходимых для роста нормальных тканей.
Опухоли можно поддерживать в течение многих лет in vitro, и при их использовании они способны вызывать опухоли у здоровых растений. В природных условиях корончатые галлы образуются в месте соединения корня со стеблем (у корневой шейки), откуда и произошло их название корончатый галл. Однако корончатые галлы могут развиваться и на подземных частях растения, например на корнях плодовых деревьев, и на надземных, например на стебле винограда.
В лаборатории эти заболевания можно вызвать у здоровых растений экспериментально, путем инфицирования их бактериями. Растения перед инокуляцией должны быть поранены, при этом опухоли возникают в поврежденных сайтах растения, обычно на стебле или листьях растения. Кроме целых растений в качестве тест-объектов используются экспланты, например ломтики моркови и кусочки других органов растений.
Ткани корончатых галлов содержат более высокие уровни ауксина и цитокининов. Выявлено еще одно наследуемое изменение в клетках корончатых галлов — это синтез опинов. Необычное для растений соединение, производное аргинина, обнаруженное лишь в определенных опухолевых линиях, было названо октопином. Затем было показано, что другими опухолевыми линиями синтезируется еще одно соединение — нопалин, также производное аргинина. В зависимости от типа индуцируемого в опухоли опина штаммы A. tumefaciens и находящиеся в них Ti-плазмиды получили соответствующее обозначение — октопиновые или нопалиновые.
Агробактерии, индуцирующие опухоли, в которых не обнаруживается ни нопалин, ни октопин, ранее обозначались как штаммы нулевого типа. Позднее было показано, что в опухолях нулевого типа синтезируются опины третьего класса — агропины. Обнаружены также и другие типы опинов. Поскольку все опины обнаруживаются только в опухолевых клетках и отсутствуют в клетках нормальных растений или клетках растительных опухолей других типов, то опины могут рассматриваться как специфические биохимические маркеры для клеток корончатых галлов.
Опухоли, развивающиеся из одной или нескольких клеток, быстро разрастаются в крупные образования, диаметр которых на определенных видах деревьев может достигать одного метра. Типичная неорганизованная опухоль представляет собой более или менее округлую дедифференцированную массу клеток (каллус), которая может иметь гладкую или шероховатую поверхность, быть паренхиматозной или одревесневшей. Иногда на периферии таких опухолей формируются листовидные структуры (тератомы), иногда — придаточные корни. Нередко на зараженных растениях наблюдаются вторичные опухоли, значительно удаленные от первичных. Обычно они обнаруживаются выше первичной опухоли, что предполагает движение бактерий или трансформирующего агента в направлении транспирации.
Распространение Agrobacterium и других фитопатогенных бактерий по межклетникам и ксилеме является хорошо доказанным фактом. Агробактерии могут передвигаться на большие дистанции со значительной скоростью. Очевидно, это не является единственной причиной индукции вторичных опухолей. Организацию опухолей, а именно форму, величину и характер развития, определяют три фактора:
• штамм агробактерий,
• генотип растения-хозяина,
• физиологическое состояние инфицируемых растительных клеток.
Agrobacterium имеет очень широкий круг растений-хозяев и может инфицировать практически все двудольные растения. Долгое время считалось, что однодольные растения не чувствительны к агробактериальной инфекции. В настоящее время показано, что при соблюдении определенных условий агробактерии могут инфицировать однодольные растения, в частности представителей таких семейств, как Amaryllidaceae, Liliaceae, Gramineae, Iridaceae и некоторых других. Однако существуют определенные вариации круга хозяев для различных штаммов Agrobacterium: некоторые штаммы способны вызывать галлообразование на отдельных видах растений, но не инфицируют другие. Различные сорта одного и того же растения также могут иметь различную чувствительность к данному бактериальному штамму.
Невозможность заражения в природе обуславливается отсутствием соответствующих рецепторов, необходимых для взаимодействия с бактериями. Другим фактором, препятствующим инфицированию однодольных агробактериями, возможно, является отсутствие в клетках растений низкомолекулярных индукторов вирулентности Agrobacterium, например ацетосирингона, которые обычно присутствуют в клеточном соке при поранении двудольных растений.
Подробная информация о структуре плазмид Agrobacterium получена путем их рестрикционного или физического картирования. В результате исследований обнаружено четыре основных области гомологии между октопиновой и нопалиновой плазмидами. Две консервативные (области А и D) вовлечены в онкогенность, еще одна (В) соответствует области контроля репликации плазмиды, в то время как последняя (С) кодирует функции конъюгативного переноса (рис. 14).
Таким образом, кроме Т-ДНК в плазмидах имеются область, кодирующая функцию конъюгации (Tra), область репликации (Ori V) и область вирулентности (Vir). Последовательности Ti-плазмиды, фланкирующие Т-ДНК (пограничные или концевые области), играют важную роль в интеграции в растительный геном и содержат несовершенные прямые повторы по 24—25 п. н. Делеция левой границы Т-ДНК не влияет на опухолеобразование, но удаление правой пограничной области приводит практически к полной утрате вирулентности. Показано, что делеция правого повтора или его части приводит к потере способности Т-ДНК включаться в растительную ДНК. Учитывая важную роль концов Т-области в переносе Т-ДНК, можно предположить, что любой сегмент ДНК, встроенный между этими концами, может быть перенесен в растения как часть Т-ДНК. Плазмиды модифицируют таким образом, чтобы удалить все онкогенные последовательности, так как они не принимают участие ни в переносе, ни в интеграции в геном клетки-хозяина. На место этих генов можно встроить чужеродную ДНК, а плазмида теряет свои онкогенные свойства. Неонкогенные Т-ДНК, присутствующие в растениях - регенерантах, передаются согласно законам Менделя. Разработаны два метода для введения Ti-плазмидных последовательностей, содержащих нужный ген, в растение.
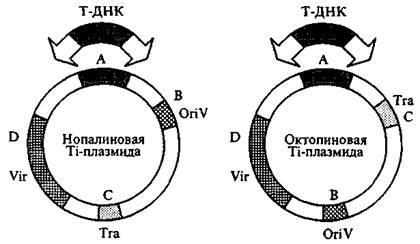
Рисунок 11. - Структура Тi-плазмид нопалинового и октопинового типа
Первый метод — метод «промежуточных векторов» (коинтегративных векторов) — основан на использовании плазмиды кишечной палочки pBR 322 (рис. 15). Т-ДНК вырезают из Ti-плазмиды с помощью рестриктаз и встраивают в плазмиду pBR 322 для клонирования в Е. соli. Бактерии, содержащие плазмиду с Т-ДНК, размножают, после чего эту плазмиду выделяют. Затем в клонированную Т-ДНК с использованием рестриктаз встраивают нужный ген. Эту рекомбинантную молекулу, содержащую Т-ДНК со встроенным в нее геном, снова размножают в большом количестве, то есть клонируют в кишечной палочке. Затем с помощью конъюгации вводят в клетки агробактерии, несущие полную Ti-плазмиду.
Между Т-сегментами нативной Ti-плазмиды и промежуточного вектора происходит гомологичная рекомбинация. В результате этого Т-ДНК со встроенным геном включается в нативную Ti-плазмиду, замещая нормальную ДНК. Получаются клетки А. tumefaciens, несущие Ti-плазмиды со встроенными в Т-сегмент нужными генами. Далее их перенос в клетки растения осуществляется обычным способом, характерным для агробактерий.
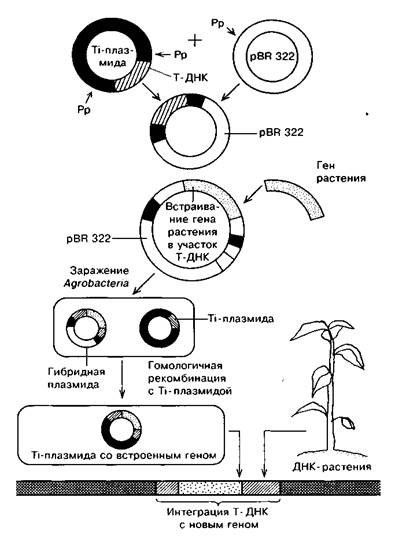
Рисунок 12. - Создание коинтегративного вектора на основе Тi-плазмиды: Рр - расщепление рестриктазой
Второй метод основан на создании системы бинарных (двойных) векторов.
Последние исследования показали, что для заражения и трансформации не нужна целая Ti-плазмида, а достаточны только пограничные области Т-ДНК и один участок Ti-плазмиды, ответственный за вирулентность. Причем эти два участка ДНК не обязательно должны находиться в одной и той же плазмиде. Если клетки агробактерий содержат Ti-плазмиду с сегментом vir и другую плазмиду с Т-ДНК, эти бактерии могут трансформировать клетки растений. При этом Т-ДНК с любыми встроенными в нее генами интегрирует с геномом растения, для этого не нужна гомологичная рекомбинация в бактериальных клетках. Для осуществления экспрессии чужеродных генов, нужен специфический промотор из Т-ДНК, например, промотор нопалинсинтетазы.
Показано, что он функционирует в клетках растений и может быть легко соединен с кодирующей последовательностью чужеродного гена в широко распространенных субклонах Ti-плазмид. Другое преимущество данного промотора заключается в том, что он функционирует в каллусах и в большинстве органов растений. Эффективность трансформации с помощью модифицированной Т-ДНК агробактерий превосходит на сегодняшний день все другие способы переноса генов в растение.
О механизмах, с помощью которых агробактерия переносит Т-ДНК ядра растений, известно очень мало: Т-сегменты ДНК октопиновых и нопалиновых плазмид встраиваются в разные, по-видимому случайные, точки хромосом хозяина, но при этом они никогда не интегрируют с ДНК митохондрий и хлоропластов.
Для введения сконструированных Ti-плазмид в растительную клетку может быть использовано несколько методов. Наиболее простой из них природный способ — это инокуляция сконструированных штаммов в поврежденные (пораненные) области растения.
Другой метод состоит в трансформации протопластов путем кокультивирования их с агробактериями Методика кокультивации может рассматриваться как индукция опухолей в искусственных условиях: вирулентные агробактерии временно совместно культивируются с протопластами. Если агробактерии добавляются к свежевыделенным или однодневным протопластам, не наблюдается ни присоединения бактерий, ни трансформации. Существенным условием для трансформации является наличие вновь образуемых клеточных стенок у 3-дневных протопластов. Это подтверждается применением ингибиторов образования клеточной стенки, которые ингибируют и присоединение бактерий. После периода кокультивации (более суток), в течение которого наступает агрегация протопластов с бактериями, свободные бактерии удаляются повторным отмыванием. Далее растительные клетки культивируются на среде с добавлением гормонов, а через 3—4 недели небольшие колонии высеваются на безгормональную среду. На этой среде выживают только колонии трансформированных клеток.
Так были получены трансформированные растения-регенеранты табака и петунии. Этот метод дает возможность существенно расширить круг хозяев агробактерий, включая виды семейства злаковых. Эффективность кокультивирования может быть повышена применением индукторов слияния клеток (ПЭГ, кальций и др.).
Трансформация протопластов может быть проведена также кокультивированием их непосредственно с Ti-плазмидами, такие опыты были проведены с протопластами петунии, табака. Очень низкая эффективность включения Т-ДНК в протопласты, наблюдавшаяся в первых экспериментах, была затем увеличена благодаря химической стимуляции (ПЭГ). Из трансформированных клеток были получены трансгенные растения. Преимуществом этого метода является то, что отпадает необходимость в промежуточных векторах. Достижения генной инженерии растений
Первые трансгенные растения (растения табака со встроенными генами из микроорганизмов) были получены в 1983 г. Первые успешные полевые испытания трансгенных растений (устойчивые к вирусной инфекции растения табака) были проведены в США уже в 1986 г.
После прохождения всех необходимых тестов на токсичность, аллергенность, мутагенность и т.д. первые трансгенные продукты появились в продаже в США в 1994 г. Это были томаты Flavr Savr с замедленным созреванием, созданные фирмой "Calgen", а также гербицид-устойчивая соя компании "Monsanto". Уже через 1-2 года биотехнологические фирмы поставили на рынок целый ряд генетически измененных растений: томатов, кукурузы, картофеля, табака, сои, рапса, кабачков, редиса, хлопчатника.
В астоящее время получением и испытанием генетически модифицированных растений занимаются сотни коммерческих фирм во всем мире с совокупным капиталом более ста миллиардов долларов. В 1999 г. трансгенные растения были высажены на общей площади порядка 40 млн. га, что превышает размеры такой страны, как Великобритания. В США генетически модифицированные растения (GM Crops) составляют сейчас около 50% посевов кукурузы и сои и более 30-40% посевов хлопчатника. Это говорит о том, что генно-инженерная биотехнология растений уже стала важной отраслью производства продовольствия и других полезных продуктов, привлекающей значительные людские ресурсы и финансовые потоки. В ближайшие годы ожидается дальнейшее быстрое увеличение площадей, занятых трансгенными формами культурных растений.
Первая волна трансгенных растений, допущенных для практического применения, содержала дополнительные гены устойчивости (к болезням, гербицидам, вредителям, порче при хранении, стрессам).
Нынешний этап развития генетической инженерии растений получил название "метаболическая инженерия". При этом ставится задача не столько улучшить те или иные имеющиеся качества растения, как при традиционной селекции, сколько научить растение производить совершенно новые соединения, используемые в медицине, химическом производстве и других областях. Этими соединениями могут быть, например, особые жирные кислоты, полезные белки с высоким содержанием незаменимых аминокислот, модифицированные полисахариды, съедобные вакцины, антитела, интерфероны и другие "лекарственные" белки, новые полимеры, не засоряющие окружающую среду и многое, многое другое. Использование трансгенных растений позволяет наладить масштабное и дешевое производство таких веществ и тем самым сделать их более доступными для широкого потребления.
Улучшение качества запасных белков
Запасные белки основных культурных видов кодируются семейством близкородственных генов. Накопление запасных белков семян – сложный биосинтетический процесс. Первая генноинженерная попытка улучшения свойства одного растения путем введения гена запасного белка от другого была, проведена Д. Кемпом и Т. Холлом в 1983 г. в США. Ген фазеолина бобов с помощью Ti-плазмиды был перенесен в геном подсолнечника. Результатом этого опыта было лишь химерное растение, получившее название санбин. В клетках подсолнечника были обнаружены иммунологически родственные фазеолиновые полипептиды, что подтверждало факт переноса гена между растениями, относящимися к различным семействам
Позднее ген фазеолина был передан клеткам табака: в растениях-регенерантах ген экспрессировался во всех тканях, хотя и в малых количествах. Неспецифическая экспрессия фазеолинового гена, так же как и в случае переноса его в клетки подсолнечника, сильно отличается от экспрессии этого гена в зрелых семядолях бобов где фазеолин составлял 25—50% от общего белка. Этот факт указывает на необходимость сохранения и других регуляторных сигналов этого гена при конструировании химерных растений и на важность контроля экспрессии генов в процессе онтогенеза растений.
Ген, кодирующий запасной белок кукурузы – зеин, после интеграции его в Т-ДНК был перенесен в геном подсолнечника следующим образом. Штаммы агробактерий, содержащие Ti-плазмиды с геном зеина, использовали для индукции опухолей в стеблях подсолнечника. Некоторые из полученных опухолей содержали мРНК, синтезируемые с генов кукурузы, что дает основание рассматривать эти результаты как первое доказательство транскрипции гена однодольного растения в двудольном. Однако присутствие зеинового белка в тканях подсолнечника не обнаружилось.
Более реальной задачей для генетической инженерии считается улучшение аминокислотного состава белков. Как известно, в запасном белке большинства злаковых наблюдается дефицит лизина, треонина, триптофана, у бобовых — метионина и цистеина. Введение в эти белки дополнительных количеств дефицитных аминокислот могло бы ликвидировать аминокислотный дисбаланс. Методами традиционной селекции удалось существенно повысить содержание лизина в запасных белках злаковых. Во всех этих случаях часть проламинов (спирторастворимые запасные белки злаковых) заменялась другими белками, содержащими много лизина. Однако у таких растении уменьшались размеры зерна и снижалась урожайность. По-видимому, проламины необходимы для формирования нормального зерна, и их замена другими белками отрицательно влияет на урожайность. Учитывая это обстоятельство, для улучшения качества запасного белка зерновых нужен такой белок, который не только отличался бы высоким содержанием лизина и треонина, но и мог полноценно заменить определенную часть проламинов при формировании зерна.
Растения могут производить и белки животного происхождения. Так, встраивание в геном растений Arabidopsis thaliana и Brassica napus химерного гена, состоящего из части гена запасного 25-белка арабидопсиса и кодирующей части для нейропептида — энкефалина, приводило к синтезу химерного белка до 200 нг на 1 г семени. Два структурных белковых домена были связаны последовательностью, узнаваемой трипсином, что давало возможность в дальнейшем легко изолировать чистый энкефалин.
В другом эксперименте удалось после скрещивания трансгенных растений, в одном из которых был встроен ген гамма-субъединицы, а во втором - ген каппа-субъединицы иммуноглобулина, получить у потомства экспрессию обеих цепей. В результате растение формировало антитела, составляющие до 1,3% суммарного белка листьев. Также было показано, что в растениях табака могут собираться полностью функциональные секреторные моноклональные иммуноглобулины. Секреторные иммуноглобулины обычно выделяются в ротовую полость и желудок человека и животных и служат первым барьером на пути кишечных инфекций. В упомянутой выше работе получили продукцию в растениях моноклональных антител, которые были специфичны для Streptococcus mutans - бактерий, вызывающих зубной кариес. Предполагается, что на основе таких моноклональных антител, продуцируемых трансгенными растениями, удастся создать действительно антикариесную зубную пасту. Из других белков животного происхождения, которые представляют интерес для медицины, показана продукция в растениях человеческого β-интерферона.
Разработаны также подходы, позволяющие получать бактериальные антигены в растениях и использовать их в качестве вакцин. Получен картофель, экспрессирующий олигомеры нетоксичной субъединицы β-токсина холеры. Эти трансгенные растения могут быть использованы для получения дешевой вакцины от холеры.
Жиры
Важнейшим сырьем для получения разного рода химических веществ являются жирные кислоты — основной компонент растительного масла. По своей структуре это углеродные цепи, которые обладают различными физико-химическими свойствами в зависимости от своей длины и степени насыщения углеродных связей. В 1995 году была закончена экспериментальная проверка и получено разрешение от федеральных властей США на выращивание и коммерческое использование трансгенных растений рапса с измененным составом растительного масла, включающего вместе с обычными 16- и 18-членными жирными кислотами также и до 45% 12-членной жирной кислоты - лаурата. Это вещество широко используется для производства стиральных порошков, шампуней, косметики.
Экспериментальная работа заключалась в том, что был клонирован ген специфической тиоэстеразы из растения Umbellularia califomica, где содержание лаурата в жире семян достигало 70%. Структурная часть гена этого фермента под контролем промотора-терминатора гена белка, специфического для ранней стадии семяобразования, была встроена в геном рапса и арабидопсиса, что и привело к увеличению содержания лаурата в масле этих растений.
Из других проектов, связанных с изменением состава жирных кислот, можно упомянуть работы, ставящие целью повышение или снижение содержания ненасыщенных жирных кислот в растительном масле. Интересными представляются эксперименты с петрозелиновой кислотой — изомером олеиновой кислоты, где двойная связь находится за шестым углеродным членом. Эта жирная кислота входит в состав масла кориандра и определяет его более высокую температуру плавления (33°С), в то время как при наличии олеиновой кислоты температура плавления составляет только 12°С. Предполагается, что после переноса генов, определяющих синтез петрозелиновой кислоты, в растения - продуценты растительного масла удастся производить диетический маргарин, содержащий ненасыщенную жирную кислоту. Кроме того, из петрозелиновой кислоты очень легко получать лаурат путем окисления озоном. Дальнейшее изучение специфики биохимического синтеза жирных кислот, по-видимому, приведет к возможности управлять этим синтезом с целью получения жирных кислот различной длины и различной степени насыщения, что позволит значительно изменить производство детергентов, косметики, кондитерских изделий, затвердителей, смазочных материалов, лекарств, полимеров, дизельного топлива и многого другого, что связано с использованием углеводородного сырья.
Полисахариды
Проводится работа по созданию трансгенных растений картофеля и других крахмалнакапливающих культур, в которых это вещество будет находиться в основном в виде амилопектина, то есть разветвленной форме крахмала, или же в основном только в виде амилозы, то есть линейных форм крахмала. Раствор амилопектина в воде более жидкий и прозрачный, чем у амилозы, которая при взаимодействии с водой образует ригидный гель. Так, например, крахмал, состоящий в основном из амилопектина, по-видимому, будет иметь спрос на рынке производителей различных питательных смесей, где сейчас в качестве наполнителя используется модифицированный крахмал. Генетической модификации могут подвергаться также геномы пластид и митохондрий. Такие системы позволяют значительно увеличить содержание продукта в трансгенном материале.
Создание гербицидоустойчивых растений
В новых, интенсивных сельскохозяйственных технологиях гербициды применяются очень широко. Это связано с тем. что на смену прежним экологически опасным гербицидам широкого спектра действия, обладающим токсичностью для млекопитающих и длительно сохраняющимся во внешней среде, приходят новые, более совершенные и безопасные соединения. Однако они обладают недостатком — подавляют рост не только сорняков, но и культурных растений Такие высокоэффективные гербициды, как, глифосат, атразины интенсивно изучаются на предмет выявления механизма толерантности к ним некоторых сорняков. Так, на полях, где широко используют атразин, довольно часто появляются атразинустойчивые биотипы у многих видов растении.
Изучение механизма устойчивости к гербицидам с целью получения методами генетической инженерии культурных растений, обладающих этим признаком, включает следующие этапы: выявление биохимических мишеней действия гербицидов в растительной клетке: отбор устойчивых к данному гербициду организмов в качестве источников генов устойчивости: клонирование этих генов: введение их в культурные растения и изучение их функционирования
Существуют четыре принципиально различных механизма, которые могут обеспечивать устойчивость к тем или иным химическим соединениям, включая гербициды: транспортный, элиминирующий, регуляционный и контактный. Транспортный механизм устойчивости заключается в невозможности проникновения гербицида в клетку. При действии элиминирующего механизма устойчивости вещества, попавшие внутрь клетки, могут разрушаться с помощью индуцируемых клеточных факторов, чаще всего деградирующих ферментов, а также подвергаться тому или иному виду модификации, образуя неактивные безвредные для клетки продукты. При регуляционной резистентности белок или фермент клетки, инактивирующийся под действием гербицида, начинает усиленно синтезироваться, ликвидируя таким образом дефицит нужного метаболита в клетке. Контактный механизм устойчивости обеспечивается изменением структуры мишени (белок или фермент), взаимодействием с которым связано повреждающее действие гербицида
Установлено, что признак гербицидоустойчивости является моногенным, то есть признак детерминируется чаще всего одним-единственным геном. Это очень облегчает возможность использования технологии рекомбинантной ДНК для передачи этого признака. Гены, кодирующие те или иные ферменты деструкции и модификации гербицидов, могут быть с успехом использованы для создания гербицидоустойчивых растении методами генетической инженерии.
Традиционные методы селекции создания сортов, устойчивых к гербицидам, очень, длительны и малорезультативны. Наиболее широко применяемый за рубежом гербицид глифосат (коммерческое название Roundup) подавляет синтез важнейших ароматических аминокислот, воздействуя на фермент 5-енолпирувилшикимат-З-фосфатсинтазу (ЕПШФ-синтаза). Известные случаи устойчивости к этому гербициду связаны либо с повышением уровня синтеза этого фермента (регуляционный механизм), либо с возникновением мутантного фермента, нечувствительного к глифосфату (контактный механизм). Из устойчивых к глифосфату растений был выделен ген ЕПШФ-синтазы и поставлен под промотор вируса мозаики цветной капусты. С помощью Ti-плазмиды эта генетическая конструкция была введена в клетки петунии. При наличии одной копии гена в регенерированных из трансформированных клеток растениях синтезировалось фермента в 20 - 40 раз больше, чем в исходных растениях, но устойчивость к глифосфату увеличилась только в 10 раз.
К числу наиболее распространенных гербицидов, используемых при обработке зерновых культур, относится атразин. Он подавляет фотосинтез, связываясь с одним из белков фотосистемы II и прекращая транспорт электронов. Устойчивость к гербициду возникает в результате точечных мутаций в этом пластохинон связывающем белке (замена серина на глицин), вследствие чего он теряет способность взаимодействовать с гербицидом. В ряде случаев удалось осуществить перенос гена мутантного белка в чувствительные к атразину растения с помощью Ti-плазмиды. Интегрированный в хромосому растений ген устойчивости был снабжен сигнальной последовательностью, которая обеспечивала транспорт синтезируемого белка в хлоропласты. Химерные растения проявляли значительную устойчивость к таким концентрациям атразина, которые вызывали гибель контрольных растений с геном белка дикого типа. Некоторые растения способны инактивировать атразин путем отщепления остатка хлора ферментом глутатион-S-трансфераза. Этот же фермент инактивирует и другие родственные гербициды триазинового ряда (пропазин, симазин и др.).
Существуют растения, естественная устойчивость которых к гербицидам основана на детоксикации. Так, устойчивость растений к хлорсульфурону может быть связана с дезактивацией молекулы гербицида путем его гидроксилирования и последующего гликозилирования введенной гидроксильной группы. Создание растений, устойчивых к патогенам и вредителям Устойчивость растений к тем или иным патогенам чаще всего является сложным мультигенным признаком.
Одновременная передача нескольких локусов трудна даже методами генной инженерии, не говоря о классических методах селекции. Более простым является другой путь. Известно, что у устойчивых растений при атаке патогенов изменяется метаболизм. Накапливаются такие соединения, как Н2О2, салициловая кислота, фитоаллексины. Повышенный уровень этих соединений способствует противостоянию растения в борьбе с патогенами.
Вот один из примеров, доказывающий роль салициловой кислоты в иммунном ответе растений. Трансгенные растения табака, которые содержат бактериальный ген, контролирующий синтез салицилат гидролазы (этот фермент разрушает салициловую кислоту), были неспособны к иммунному ответу. Поэтому изменение генно-инженерным путем уровня салициловой кислоты или выработки в растениях в ответ на патоген Н2О2 может быть перспективным для создания устойчивых трансгенных растений.
В фитовирусологии широко известен феномен индуцированной перекрестной устойчивости растений к вирусным инфекциям. Сущность этого явления состоит в том, что заражение растения одним штаммом вируса предотвращает последующую инфекцию этих растений другим вирусным штаммом. Молекулярный механизм подавления вирусной инфекции пока неясен. Показано, что для иммунизации растений достаточно введения отдельных вирусных генов, например генов капсидных белков. Так, ген белка оболочки вируса табачной мозаики перенесли в клетки табака и получили трансгенные растения, у которых 0,1% всех белков листьев был представлен вирусным белком. Значительная часть этих растений при инфицировании вирусом не проявляла никаких симптомов заболевания. Возможно, что синтезирующийся в клетках белок оболочки вируса мешает вирусной РНК нормально функционировать и формировать полноценные вирусные частицы. Установлено, что экспрессия капсидного белка вируса табачной мозаики, вируса мозаики люцерны, вируса огуречной мозаики, Х-вируса картофеля в соответствующих трансгенных растениях (табак, томаты, картофель, огурцы, перцы) обеспечивает высокий уровень их защиты от последующей вирусной инфекции. Причем у трансформированных растений не отмечалось снижения фертильности, нежелательного изменения ростовых и физиологических характеристик исходных экземпляров и их потомства. Полагают, что индуцированная устойчивость растений к вирусам обусловлена особым антивирусным белком, очень похожим на интерферон животных. Представляется возможным методом генетической инженерии усилить экспрессию гена, кодирующего этот белок, путем его амплификации или подстановки под более сильный промотор.
Следует отметить, что использование генетической инженерии для защиты растений от различных патогенных микроорганизмов в значительной мере сдерживается недостаточностью знаний о механизмах защитных реакций растений. Для борьбы с насекомыми-вредителями в растениеводстве используются химические средства — инсектициды. Однако они оказывают вредное влияние на млекопитающих, убивают и полезных насекомых, загрязняют окружающую среду, дороги, и кроме того, насекомые довольно скоро приспосабливаются к ним. Известно более 400 видов насекомых, устойчивых к используемым инсектицидам. Поэтому все большее внимание привлекают биологические средства борьбы, обеспечивающие строгую избирательность действия и отсутствие адаптации вредителей к применяемому биопестициду.
Уже довольно давно известна бактерия Bacillus thuringiensis, продуцирующая белок, являющийся очень токсичным для многих видов насекомых, в то же время безопасный для млекопитающих. Белок (дельта-эндотоксин, CRY-белок) продуцируется различными штаммами В. thuringiensis. Взаимодействие токсина с рецепторами строго специфично, что усложняет подбор комбинации токсин—насекомое. В природе найдено большое количество штаммов В. thuringiensis, чьи токсины действуют только на определенные виды насекомых. Препараты В. thuringiensis в течение десятилетий использовали для контроля насекомых на полях. Безопасность токсина и его составных белков для человека и других млекопитающих полностью доказана. Встраивание гена этого белка в геном растений дает возможность получить трансгенные растения, не поедаемые насекомыми.
Кроме видоспецифичности по действию на насекомых встраивание прокариотических генов дельта-токсинов в геном растений даже под контролем сильных эукариотических промоторов не привело к высокому уровню экспрессии. Предположительно такое явление возникло в связи с тем, что эти бактериальные гены содержат значительно больше адениновых и тиминовых нуклеотидных оснований, чем растительная ДНК. Эта проблема была решена путем создания модифицированных генов, где из природного гена вырезали и добавляли те или иные фрагменты с сохранением доменов, кодирующих активные части дельта-токсина. Так, например, с помощью таких подходов был получен картофель, устойчивый к колорадскому жуку. Получены трансгенные растения табака, способные синтезировать токсин. Такие растения были нечувствительны к гусеницам Manduca sexta. Последние погибали в течение 3 суток контакта с токсинпродуцирующими растениями. Токсинообразование и обусловленная им устойчивость к насекомым передавалась по наследству как доминантный признак.
В настоящее время так называемые Bt-растения (от В. thuringiensis) хлопка и кукурузы занимают основную долю в общем объеме генетически модифицированных растений этих культур, которые выращивают на полях США.
В связи с возможностями генной инженерии конструировать энтомопатогенные растения на основе токсина микробного происхождения еще больший интерес к себе вызывают токсины растительного происхождения. Фитотоксины являются ингибиторами белкового синтеза и осуществляют защитную функцию, направленную против насекомых-вредителей микроорганизмов и вирусов. Лучше всех среди них изучен рицин, синтезируемый в клещевине: его ген клонирован и установлена нуклеотидная последовательность. Однако высокая токсичность рицина для млекопитающих ограничивает генноинженерные работы с ним только техническими культурами, не используемыми в пищу человека и на корм животным. Токсин, вырабатываемый фитолаккой американской, эффективен против вирусов и безвреден для животных. Механизм его действия заключается в инактивации собственных рибосом при проникновении в клетки различного рода патогенов, в том числе фитовирусов. Пораженные клетки некротизируются, предотвращая размножение патогена и его распространение по растению. В настоящее время проводятся исследования по изучению гена этого белка и передаче его в другие растения.
Вирусные болезни широко распространены среди насекомых, поэтому для борьбы с насекомыми-вредителями можно использовать природные вирусы насекомых, препараты которых называют вирусными пестицидами. В отличие от ядохимикатов они обладают узким спектром действия, не убивают полезных насекомых, они быстро разрушаются во внешней среде и не опасны для растений и животных. Наряду с вирусами насекомых используются как биопестициды некоторые грибы, поражающие насекомых-вредителей. Применяемые сейчас биопестициды являются природными штаммами энтомопатогенных вирусов и грибов, однако не исключена возможность создания в будущем методами генетической инженерии новых эффективных биопестицидов.
Повышение устойчивости растений к стрессовым условиям
Растения очень часто подвергаются воздействию различных неблагоприятных факторов окружающей среды: высокие и низкие температуры, недостаток влаги, засоление почв и загазованность среды, недостаток или, напротив, избыток некоторых минеральных веществ и т. д. Этих факторов множество, поэтому и способы защиты от них многообразны — от физиологических свойств до структурных приспособлений, позволяющих преодолевать их пагубное действие.
Устойчивость растений к тому или иному стрессовому фактору является результатом воздействия множества разных генов, поэтому говорить о полной передаче признаков толерантности от одного вида растения другому генноинженерными методами не приходится. Тем не менее у генетической инженерии имеются определенные возможности для повышения устойчивости растений. Это касается работы с отдельными генами, контролирующими метаболические ответы растений на стрессовые условия, например сверхпродукцию пролина в ответ на осмотический шок, на действие засоления, синтез особых белков в ответ на тепловой шок и т. д. Дальнейшее углубленное изучение физиологической, биохимической и генетической основы ответной реакции растения на условия среды, несомненно, позволит применять методы генетической инженерии для конструирования устойчивых растений.
Пока можно отметить лишь косвенный подход для получения морозоустойчивых растений, основанный на генноинженерных манипуляциях с Pseudomonas syringae. Этот микроорганизм, сосуществующий с растениями, способствует их повреждению ранними заморозками Механизм явления связан с тем, что клетки микроорганизма синтезируют особый белок, локализующийся во внешней мембране и являющийся центром кристаллизации льда. Известно, что формирование льда в воде зависит от веществ, могущих служить центрами образования льда. Белок, вызывающий формирование кристаллов льда в различных частях растения (листья, стебли, корни), является одним из главных факторов, ответственных за повреждение тканей растений, чувствительных к ранним заморозкам. Многочисленные эксперименты в строго контролируемых условиях показали, что стерильные растения не повреждались заморозками вплоть до —6—8° С, тогда как у растений, имеющих соответствующую микрофлору, повреждения возникали уже при температурах —1,5—2° С. Мутанты этих бактерий, потерявшие способность синтезировать белок, вызывающий формирование кристаллов льда, не повышали температуру образования льда, и растения с такой микрофлорой были устойчивы к заморозкам. Штамм таких бактерий, распыленный над клубнями картофеля, конкурировал с обычными бактериями, что приводило к повышению морозоустойчивости растений. Возможно, такие бактерии, созданные с помощью методов генной инженерии и используемые в качестве компонента внешней среды, будут служить для борьбы с заморозками.
Повышение эффективности биологической азотфиксации
Хорошо изучен фермент ответственный за восстановление молекулярного азота до аммония. — нитрогеназа. Структура нитрогеназы одинакова у всех азотфиксирующих организмов. При фиксации азота непременным физиологическим условием является защита нитрогеназы от разрушения под действием кислорода. Лучше всех среди азотфиксаторов изучены ризобии, образующие симбиоз с бобовыми растениями, и свободноживущая бактерия Klebsiella pneumoniae. Установлено, что у этих бактерий за фиксацию азота ответственно 17 генов — так называемых nif-генов. Все эти гены сцеплены друг с другом и расположены в хромосоме между генами ферментов биосинтеза гистидина и генами, определяющими усвоение шикимовой кислоты. У быстрорастущей ризобии nif-гены существуют в форме мегаплазмиды, содержащей 200—300 тысяч пар нуклеотидов.
Среди генов азотфиксации выявлены гены, контролирующие структуру нитрогеназы, белковый фактор, принимающий участие в транспорте электронов, регуляторные гены. Регуляция генов азотфиксации довольно сложна, поэтому генноинженерный перенос азотфиксирующей функции от бактерий непосредственно высшим растениям в настоящее время уже не обсуждается. Как показали эксперименты, даже в самом простом эукариотическом организме — дрожжах не удалось добиться экспрессии nif-генов, хотя они и сохранялись в течение 50 генераций.
Эти опыты показали, что диазотрофность (азот-фиксация) свойственна исключительно прокариотическим организмам, и nif-гены не смогли преодолеть барьер, разделяющий прокариоты и эукариоты, из-за слишком сложной своей структуры и регуляции генами, расположенными вне nif-области. Возможно, более удачным окажется перенос nif-генов с помощью Ti-плазмид в хлоропласты, поскольку механизмы экспрессии генов в хлоропластах и в клетках прокариот близки. В любом случае нитрогеназа должна быть защищена от ингибирующего действия кислорода. Кроме того, фиксация атмосферного азота — очень энергоемкий процесс. Вряд ли растение под влиянием nif-генов может так кардинально изменить свой метаболизм, чтобы создать все эти условия. Хотя не исключено, что в будущем методами генетической инженерии можно будет создать более экономно работающий нитрогеназный комплекс.
Более реально использование генноинженерных методов для решения следующих задач: повышение способности ризобии колонизировать бобовые растения, повышение эффективности фиксации и ассимиляции азота путем воздействия на генетический механизм, создание новых азотфиксирующих микроорганизмов путем введения в них nif-генов, передача способности к симбиозу от бобовых растений к другим.
Первостепенной задачей генетической инженерии для повышения эффективности биологической фиксации азота является создание штаммов ризобии с усиленной азотфиксацией и колонизирующей способностью. Колонизация бобовых растений ризобиями протекает очень медленно, лишь единичные из них дают начало клубенькам. Это происходит потому, что местом инвазии ризобии является только одна небольшая область между точкой роста корня и ближайшим к ней корневым волоском, находящимся на стадии формирования. Все остальные части корня и развившиеся корневые волоски растения нечувствительны к колонизации. В ряде случаев сформировавшиеся клубеньки оказываются неспособными фиксировать азот, что зависит от многих растительных генов (выявлено не менее пяти), в частности от неблагоприятного сочетания двух рецессивных генов.
Традиционными методами генетики и селекции удалось получить лабораторные штаммы ризобий с более высокой колонизирующей способностью. Но они в полевых условиях испытывают конкуренцию со стороны местных штаммов. Повышение их конкурентоспособности, видимо, можно осуществить генноинженерными методами. Повышение эффективности процесса азотфиксации возможно применением генноинженерных приемов, основанных на увеличении копий гена, усилении транскрипции тех генов, продукты которых образуют «узкое» место в каскадном механизме азотфиксации, путем введения более сильных промоторов и т. п. Важно повышение коэффициента полезного действия самой нитро-геназной системы, осуществляющей непосредственное восстановление молекулярного азота в аммиак.
Повышение эффективности фотосинтеза
С4-растения характеризуются высокими темпами роста и скоростью фотосинтеза, у них практически отсутствует видимое фотодыхание. У большинства сельскохозяйственных культур, относящихся к С3-растениям, высокая интенсивность фотодыхания. Фотосинтез и фотодыхание — тесно связанные процессы, в основе которых лежит бифункциональная активность одного и того же ключевого фермента — рибулозобисфосфат-карбоксилазы (РуБФК). РуБФ-карбоксилаза может присоединять не только С02, но и 02, то есть осуществляет реакции карбоксилирования и оксигенирования. При оксигенировании РуБФ образуется фосфогликолат, который служит основным субстратом фотодыхания — процесса выброса С02 на свету, в результате чего теряется часть фотосинтетических продуктов. Низкое фотодыхание у С4-растений объясняется не отсутствием ферментов гликолатного пути, а ограничением оксигеназной реакции, а также реассимиляцией С02 фотодыхания.
Одной из задач, стоящих перед генетической инженерией, является исследование возможности создания РуБФК с преобладающей карбоксилазной активностью.
Получение растений с новыми свойствам
В последние годы ученые используют новый подход для получения трансгенных растений с "antisense RNA" (перевернутой или антисмысловой РНК), который позволяет управлять работой интересуемого гена. В этом случае при конструировании вектора копию ДНК (к-ДНК) встраиваемого гена переворачивают на 180°. В результате в трансгенном растении образуется нормальная молекула мРНК и перевернутая, которая в силу комплементарности нормальной мРНК образует с ней комплекс и закодированный белок не синтезируется.
Такой подход использован для получения трансгенных растений томатов с улучшенным качеством плодов. Вектор включал к-ДНК гена PG, контролирующего синтез полигалактуроназы - фермента, участвующего в разрушении пектина, основного компонента межклеточного пространства растительных тканей. Продукт гена PG синтезируется в период созревания плодов томатов, а увеличение его количества приводит к тому, что томаты становятся более мягкими, что значительно сокращает срок их хранения. Отключение этого гена в трансгенах позволило получить растения томатов с новыми свойствами плодов, которые не только значительно дольше сохранялись, но и сами растения были более устойчивы к грибным заболеваниям.
Такой же подход можно применить для регулирования сроков созревания томатов, а в качестве мишени в этом случае используют ген EFE (ethylene-forming enzyme), продуктом которого является фермент, участвующий в биосинтезе этилена. Этилен - это газообразный гормон, одной из функций которого является контроль за процессом созревания плодов.
Cтратегия антисмысловых конструкций широко применима для модификации экспрессии генов. Эта стратегия используется не только для получения растений с новыми качествами, но и для фундаментальных исследований в генетике растений. Следует упомянуть еще об одном направлении в генной инженерии растений, которое до недавнего времени в основном использовали в фундаментальных исследованиях - для изучения роли гормонов в развитии растений. Суть экспериментов заключалась в получении трансгенных растений с комбинацией определенных бактериальных гормональных генов, например только iaaM или ipt т.д. Эти эксперименты внесли существенный вклад в доказательство роли ауксинов и цитокининов в дифференцировке растений.
В последние годы этот подход стали использовать в практической селекции. Оказалось, что плоды трансгенных растений с геном iaaM, находящимся под промотором гена Def (ген, который экспрессируется только в плодах), являются партенокарпическими, то есть сформировавшимися без опыления. Партенокарпические плоды характеризуются либо полным отсутствием семян, либо очень небольшим их количеством, что позволяет решить проблему "лишних косточек", например в арбузе, цитрусовых и т.д. Уже получены трансгенные растения кабачков, которые в целом не отличаются от контрольных, но практически не содержат семян.
Обезоруженную, лишенную онкогенов Ti-плазмиду ученые активно используют для получения мутаций. Этот метод носит название Т-ДНК-инсерционного мутагенеза. Т-ДНК, встраиваясь в геном растения, выключает ген, в который она встроилась, а по утрате функции можно легко отбирать мутанты (явление сайлесинга – замолкания генов). Этот метод замечателен также тем, что позволяет сразу обнаружить и клонировать соответствующий ген. В настоящее время таким способом получено множество новых мутаций растений и соответствующие гены клонированы. М. А. Раменской на основе Т-ДНК мутагенеза получены растения томатов с неспецифической устойчивостью к фитофторозу. Не менее интере сен и другой аспект работ - получены трансгенные растения с измененными декоративными свойствами. Один из примеров - это получение растений петунии с разноцветными цветками. На очереди голубые розы с геном, контролирующим синтез голубого пигмента, клонированным из дельфиниума. Проблемы биобезопасности трансгенных растений
Одним из главных возражений против употребления "трансгенных" продуктов питания является наличие во многих из них генов устойчивости к антибиотику (в частности, к канамицину), которые содержались в исходной конструкции ДНК в качестве селективных.
Предполагается, что эти гены устойчивости могут при переваривании пищи передаваться эндогенной микрофлоре, в том числе патогенной, в результате чего микробы могут приобрести резистентность к данному антибиотику. Однако в реальности вероятность такого события ничтожно мала - многочисленные эксперименты и наблюдения в природе относительно подобного горизонтального переноса генов до сих пор давали только отрицательные результаты.
Не стоит забывать, что встраиваемые в растения гены устойчивости "настроены" для экспрессии лишь в эукариотических, но не бактериальных клетках. Надо учесть и то, что эти селективные гены взяты из природных популяций микроорганизмов, где они сейчас широко распространены в результате активного применения антибиотиков в медицинской практике. Поэтому вероятность попадания гена устойчивости к антибиотику в микрофлору человека из природного резервуара несравнимо реальнее, чем при употреблении трансгенных растений. Однако, учитывая настроения общественности, разрабатываются подходы, для исключения присутствия "подозрительных" генов в коммерциализированных трансгенных формах.
В большинстве случаев маркерные гены устойчивости к антибиотикам сейчас заменяют на гены устойчивости к гербицидам. Правда, применение "гербицидных" генов также встречает возражения, но уже защитников окружающей среды. Предложено несколько способов избирательной элиминации маркерного гена после получения желаемого трансгенного растения, когда он фактически уже не нужен.
Очень перспективным представляется замена селективных генов на репортерные при отборе трансгенных форм растений, либо использование альтернативных селективных генов, таких как гены синтеза фитогормонов или гидролиза особых форм полисахаридов при выращивании растений в культуральной среде. Таким образом, даже эта виртуальная опасность, связанная с генами устойчивости к антибиотику, в скором времени перестанет существовать.
Что касается возможной токсичности или аллергенности трансгенных растений, то здесь применяют те же жесткие стандарты, как и для полученных традиционным путем новых сортов культурных растений или новых видов продуктов питания. Никаких особых отличий трансгенных растений от обычных по этим параметрам ожидать не приходится (разве что в лучшую сторону при блокировании синтеза токсинов или аллергенов), да и действительно, как правило, не наблюдается на практике.
Проблема возможного ущерба для окружающей среды имеет несколько аспектов. Во-первых, существует опасение, что устойчивые к гербицидам культурные растения могут при межвидовом опылении передавать эти гены близкородственным сорнякам, которые могут превратиться в неистребимые суперсорняки (superweeds). Хотя вероятность такого нежелательного развития событий для большинства сельскохозяйственных культур очень мала, генные инженеры и ученые-аграрии активно разрабатывают подходы для исключения подобной опасности. Здесь, правда, надо отметить, что данный вопрос также не нов, так как в практике сельского хозяйства уже давно используется ряд устойчивых к гербицидам сортов, полученных путем обычной селекции. При этом никакой экологической катастрофы широкое использование таких устойчивых сортов до сих пор не вызвало.
Тем не менее и в этом случае, чтобы отвести любые возражения от трансгенных растений, пробуют, например, вводить в растения не один, а сразу несколько генов устойчивости к разным гербицидам. Передача нескольких генов сорнякам гораздо менее вероятна, чем одного гена. Кроме того, мультигербицидная устойчивость позволит чередовать разные гербициды при обработке посевов, что не даст возможности для распространения какого-либо определенного гена устойчивости в сорняках.
Предлагается также вводить гены устойчивости не в ядерный, а в хлоропластный геном. Это может предотвратить нежелательный дрейф генов с помощью пыльцы, так как хлоропласты наследуются только по материнской линии.
Еще один генно-инженерный путь борьбы с сорняками без использования генов резистентности к гербицидам вообще – биотрансгенный. Речь идет об использовании мелких животных, например, кроликов, для поедания сорняков на полях. При этом, чтобы оградить от поедания культурные растения, в них можно ввести какой-либо ген, делающий их непривлекательными (запах, вкус) для данного животного. Такой биотрансгенный подход сразу снял бы большинство выдвигаемых сейчас возражений против трансгенных культур.
Близкие по сути экологические возражения касаются трансгенных растений со встроенными "инсектицидными" генами, способных, как считают, спровоцировать у насекомых-вредителей возникновение массовой резистентности. Здесь также предложены действенные способы для уменьшения этой опасности, например, использование генов нескольких разных токсинов и/или индуцибельных промоторов, быстро активирующихся при нападении насекомых на растение. Данная проблема в общем не нова, так как многие из инсектицидов, используемых сейчас на "генном уровне", давно применяют в виде чистого вещества для опрыскивания посевов.
Еще одно нежелательное следствие использования трансгенных растений с генами инсектицидов заключается в том, что пыльца этих растений может быть токсичной и для полезных насекомых, которые данной пыльцой питаются. Некоторые экспериментальные данные говорят о том. что такая опасность действительно существует, хотя о ее возможных масштабах говорить пока трудно. Однако и здесь уже предложены и испытаны адекватные генно-инженерные решения, например, использование трансгеноза через хлоропластную ДНК, или промоторов, не работающих в пыльце.
Надежды, которые возлагаютсяна генетически модифицированные (ГМ) растения, можно подразделить на два основных направления:
1.Усовершенствование качественных характеристик продукции растениеводства.
2. Увеличение продуктивности и стабильности растениеводства путем повышения резистентности растений к неблагоприятным факторам.
Создание генетически модифицированных растений чаще всего выполняется для решения следующих конкретных задач:
1) В целях увеличения урожайности путем повышения:
а) резистентности к патогенам;
б)резистентности к гербицидам;
в) устойчивости к неблагоприятным температурам, невысокому качеству почв;
г) улучшения характеристики продуктивности (вкусовых и питательных качеств, оптимальный метаболизм).
2) В фармакологических целях:
а) получение продуцентов терапевтических агентов;
б) продуцентов антигенов, обеспечения пищевой «пассивной» иммунизации.
Основные задачи ДНК –технологии в создании ГМ – растений в современных условиях развития сельского хозяйства и общества довольно многообразны и заключаются в следующем:
1. Получение гибридов (совместимость, мужская стерильность).
2. Оптимизация роста и развития растений (изменение габитуса растений – например, высоты, формы листьев и корневой системы и др.; изменения в цветении – например, строении и окраске цветков, времени зацветания).
3. Оптимизация питания растений (фиксация атмосферного азота небобовыми растениями; улучшение поглощения элементов минерального питания; повышение эффективности фотосинтеза).
4. Улучшение качества продукции (изменение состава и/или количества жиров; изменение вкуса и запаха пищевых продуктов; получение новых видов лекарственного сырья; изменение свойств волокна для текстильного сырья; изменение качества и сроков созревания или хранения плодов).
5. Повышение устойчивости к абиотическим факторам стресса (устойчивость к засухе и засолению. Жаростойкость; устойчивость к затоплению; адаптация к холоду; устойчивость к гербицидам; устойчивость к кислотности почв и алюминию; устойчивость к тяжелым металлам).
6. Увеличение устойчивости к биотическим факторам стресса (устойчивость к вредителям4 устойчивость к бактериальным, вирусным и грибным болезням).
Среди генов, определяющих устойчивость к гербицидам, уже клонированы гены устойчивости к таким гербицидам, как глифосат (Раундап). Фосфинотрицин (Биалафос), глифосинат аммония (Баста), сульфонилмочевинные и имидозолиновые препараты. С использованием этих генов уже получены трансгенные соя, кукуруза, хлопчатник и т.д. В России также проходят испытания трансгенные культуры, устойчивые к гербицидам. В Центре «Биоинженерия» создан сорт картофеля, устойчивый к Басте, проходящий в настоящее время полевые испытания.
n Общая площадь выращивания генетически модифицированных (ГМ) трансгенных растений в 2004 г в мире составила 81 млн.га
n В основном, это ГМ, модифицированные в отношении устойчивости к патогенным агентам и гербицидам
Дата добавления: 2015-07-14; просмотров: 6244;