Химический состав мембран
Исследования показали, что основными химическими компонентами мембран являются липиды ( 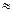 30%), белки ( 60%) и углеводы ( 10%). Минорными компонентами мембран являются нуклеиновые кислоты, полиамины, неорганические ионы.
30%), белки ( 60%) и углеводы ( 10%). Минорными компонентами мембран являются нуклеиновые кислоты, полиамины, неорганические ионы.
Липиды, входящие в состав мембран, принадлежат к трем классам. Это фосфолипиды, гликолипиды и стероиды.
Фосфолипидыпостроены по единому плану, их молекула имеет полярную головку и два неполярных хвоста.
В состав головки фосфолипида обязательно входят:
А) остаток спирта (реже аминокислоты);
Б) остаток фосфорной кислоты;
В) по вариантам, либо:
- глицерин (глицерофосфолипиды),
- многоатомный спирт сфингозин (сфингофосфолипиды).
Неполярные хвосты всех молекул фосфолипидов образуются жирными кислотами, определяющими разнообразие их представительства.
Глицерофосфолипиды мембран растительных клеток содержат в основном пальмитиновую (С16:1 (9)), олеиновую (С18:1(9)), линолевую (С18:2(9,12)) жирные кислоты. Жирные кислоты с количеством атомов углерода более 20 – 24 в растительных клетках встречаются редко.
Глицерофосфолипиды мембран животных клеток содержат пальмитиновую, олеиновую, стеариновую (С18:0) жирные кислоты, а также кислоты (арахидоновая и др.) с числом атомов углерода, превышающим 20.
Обычно из двух жирных кислот, образующих гидрофобный хвостглицерофосфолипида, одна ненасыщенная. Она образует связь со вторым атомом углерода глицерина и одна из ее двойных связей обязательно находится на уровне 9 атома углерода, считая от головки.
Остаток спирта (аминокислоты) головки глицерофосфолипида связан сложноэфирной связью с фосфорной кислотой. Глицерофосфолипид, лишенный спирта (аминокислоты), называется фосфатидной кислотой, которая является промежуточным продуктом в биосинтезе фосфолипидов и в свободном виде практически не встречается.
В зависимости от того, какой именно спирт (аминокислота) входит в состав головки молекулы, различают несколько классов глицерофосфолипидов. Самые распространенные:
· фосфатидилхолин, содержащий спирт холин;
· фосфатидилэтаноламин, содержащий спирт этаноламин;
· фосфатидилинозитол, содержащий циклический спирт инозитол;
· фосфатидилсерин, содержащий остаток гидроксиаминокислоты – серина.
Примерами сфингофосфолипидов могут служить сфингомиелины, которые в составе полярной головки содержат фосфохолин или фосфоэтаноламин. В составе сфингофосфолипидов одна жирная кислота. Роль второго неполярного хвоста выполняют углеродные структуры многоатомного спирта сфингозина.
Таким образом, фосфолипиды содержат группировки двух видов – полярные гидрофильные головки и неполярные гидрофобные хвосты. Вследствие этого фосфолипиды обладают амфипатическими свойствами. При рН = 7 остаток фосфорной кислоты в полярной головке заряжен отрицательно.
Гликолипиды – липиды, содержащие 2 остатка жирных кислот, спирт сфингозин и остатки моно- (цереброзиды) или олигосахаров (ганглиозиды).
Стероиды содержат стероидное ядро, образованное тремя гексагональными полностью насыщенными кольцами и одним циклопентановым кольцом. Стероиды представлены в основном холестерином (в животных клетках) или ситостерином и стигмастерином (в растительных клетках).
Физиологическая роль различных классов липидов
Фосфолипиды выполняют структурную функцию, образуя липидный бислой мембраны. Кроме этого, они могут выполнять и важные физиологические функции. Так, арахидоновая кислота, состоящая из цепи с 20 атомами углерода и с 4 двойными связями, входящая в состав фосфолипидов, служит предшественником таких биологически активных веществ, как простагландины.
Гликолипиды широко представлены в различных тканях, в частности, в нервной. Они локализованы преимущественно на наружной поверхности цитоплазматической мембраны, где их углеводные компоненты входят в число других углеводов клеточной поверхности. Ганглиозиды участвуют в дифференцировке нейрональной ткани. Ганглиозиды других клеток определяют видоспецифичность и регулируют межклеточные контакты. В иммуннокомпетентных клетках они участвуют в формировании иммунной реакции. Холестерин выполняет важную роль в модификации бислоя: он, в частности, регулирует упаковку и подвижность фосфолипидов мембраны. Кроме того, холестерин служит предшественником в образовании половых гормонов, гормонов коры надпочечников, а также желчных кислот.
Липид-липидные взаимодействия. Динамика липидов в мембране
Длина растянутой углеводородной цепи жирной кислоты, состоящей из 18 атомов углерода, составляет 2 нм, на полярную головку приходится еще 0,5 – 0,7 нм, следовательно, толщина бислоя липидов должна превышать 4 нм, но в действительности толщина двойного слоя липидов оказывается равной 3,5 – 4 нм. Причиной обнаруженного феномена является рыхлое, а не упорядоченное расположения остатков жирных кислот. Раз одна из двух жирных кислот, образующих гидрофобный хвост, ненасыщенная, как правило, увеличивается вероятность образования цис-конформации.
Наличие цис-изомеров жирных кислот в составе фосфолипидов, а также высокая скорость вращения вокруг С-С связей жирно-кислотных радикалов (например, частота вращения вокруг единичной С-С связи составляет 1010, а вокруг двойной С=С связи – 108 с-1) обеспечивают неупорядоченное расположение гидрофобных хвостов. Различные конфигурации молекул жирных кислот, возникающие при поворотах вокруг С-С связей, называют ротамерами или конформерами. Процесс изменения конформации молекул за счет таких поворотов называется транс-гош-изомеризацией. Гош-конформация (“гош” – скошенный) аналогична цис-конформации. Вероятность транс-гош-перехода весьма велика и еще более возрастает при увеличении температуры. При переходе из транс- в гош-конформацию образуются складки или кинки (от англ. Kink – петля). Такого рода изменения способны захватывать целые кластеры мембранных липидов. Считается, что именно с образованием кинков связан транспорт воды и других веществ через мембраны.
Липиды способны совершать вращательные движения вокруг оси молекулы (вращательная диффузия), перемещаться вдоль одного слоя (латеральная диффузия), а также перемещаться из одного слоя в другой (трансмембранный переход или флип-флоп). Наибольшую скорость имеют вращательная и латеральная диффузия. Например, коэффициент латеральной диффузии составляет 1,8×10-8 см2 с-1. Это соответствует частоте парных перестановок соседних молекул 107 с-1. Наиболее медленным процессом является флип-флоп, что связано с высоким уровнем энергии, необходимой для проталкивания заряженных полярных головок через средний углеводородный слой мембраны. Поперечная диффузия молекул фосфолипидов на расстояние 5 нм занимает в 109 раз больше времени, чем диффузия на то же расстояние в латеральном направлении.
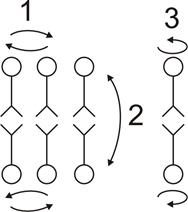
| Рис.8. Виды диффузии липидов в мембране 1-латеральная диффузия; 2-трансмембранный переход или флип-флоп; 3-вращательная диффузия. |
Сочетание быстрой диффузии молекул липидов вдоль мембраны и медленной диффузии поперек мембраны имеет большое значение для функционирования мембран. Благодаря этому поддерживается упорядоченность в молекулярной структуре мембраны, определенная ориентация белков поперек мембраны, что имеет значение для направленного переноса веществ через мембрану, кроме того, обеспечивается асимметрия бислоя липидов. Функциональная асимметрия мембраны – один из важнейших аспектов, характеризующих липид – липидные взаимодействия. Асимметрия бислоя означает, что состав липидов каждого из слоев неодинаков.
Дата добавления: 2015-06-22; просмотров: 2494;