Рост и возрастная изменчивость рыб
Познание закономерностей роста рыб, а в естественных водоемах возрастного состава стада дает представление о степени использования рыбой пищевой базы, позволяет определить время и размеры наиболее целесообразного вылова и т. д. Иначе говоря, оно дает основание для составления прогнозов о составе стада и масштабе промысла, а в прудовом рыбоводстве является основой для племенной работы, для установления наиболее выгодных с хозяйственной точки зрения сроков выращивания рыбы.
Продолжительность жизни рыб и их размеры весьма различны, но специфичны для каждого вида. Самые маленькие рыбки — бычки с Филиппинских о-вов — не превышают в длину 7,5 — 14,0мм, продолжительность их жизни — около года. В тропиках в пресных водах живут также крошечные представители семейства Cyprinodontidae (карпозубых). Другая крайность — белуга массой около 1,5т, возраст которой может составить 100 лет, или полярная акула, достигающая в длину 15 м и даже больше. Обыкновенный сом достигает массы 300 кг. 100 лет, конечно, редкость. Обычно предельный возраст ‛долгожителей‛ исчерпывается 25—30 годами. У наших пресноводных рыб предельный возраст меньше: у аральского леща — 15 лет, амурского серебряного карася — 12, амурского сазана — 16, карпа — 20 лет.
Величина рыб одного вида и одной возрастной группы из разных водоемов может различаться в зависимости от условий жизни. Например, в Подмосковье зарегистрированы сеголетки карпа массой 200 г, а на о-ве Ява — трехмесячные карпы массой 690 г.
В настоящее время в связи с интенсификацией промысла лишь ограниченное количество особей доживает до предельного видового возраста. Естественная продолжительность жизни определяется видовыми особенностями обмена веществ. Многие виды рыб погибают после первого нереста (дальневосточные лососи рода Oncorhynchus, угорь Anguilla). Некоторым видам свойственна частичная посленерестовая гибель (у благородного лосося — семги — после нереста гибнет до 87% особей, причем преимущественно самцов).
Рост неразрывно связан с развитием, являясь одной из его сторон. Специфической особенностью роста рыб является преобладание ассимиляции над диссимиляцией, благодаря чему рыба растет в течение всей жизни и после наступления половой зрелости. Характерным свойством рыб является снижение обмена (но без нарушения функций органов) при длительном недостатке или отсутствии пищи и быстрое восстановление его интенсивности при улучшении условий питания. Эта способность позволяет многим рыбам безболезненно переносить долгое зимнее голодание.
Различают весовой рост (наращивание массы тела) и линейный (увеличение длины тела). Весовой рост сильнее подвержен колебаниям в зависимости от условий питания, чем линейный, поэтому закономерности изменения роста, его специфику легче проследить на линейном росте. Вместе с тем в прудовом рыбоводстве основным показателем эффективности выращивания рыбы является весовой рост. Рыба растет неравномерно как в течение жизни, так и в течение всего года, причем в разные периоды рост ее характеризуется определенными особенностями.
Прежде всего различен характер роста рыб до и после наступления половой зрелости. Обычно до наступления половой зрелости рыбы растут наиболее быстро. Пища используется ими в основном на весовой и главным образом линейный прирост, т. е. является в основном продуцирующей. Поэтому в первые годы жизни, как правило, происходит наиболее быстрое нарастание линейных размеров. После наступления половой зрелости темп роста снижается, но наращивание массы тела продолжается и максимальное увеличение ее наблюдается именно в старшем возрасте. Значительная часть потребленной пищи расходуется на образование половых клеток и накопление резервных веществ, обеспечивающих созревание гонад, благополучный исход зимовки, миграции и т. д. Доля продуцирующей пищи уменьшается и соответственно увеличивается часть пищи, идущей на поддержание жизнедеятельности (поддерживающая пища).
В период старения организма линейный рост сильно замедляется. Пища расходуется в основном на поддержание жизненных процессов. Такова общая закономерность, отражающая характер развития рыб в течение жизни. В ее пределах рост рыб в отдельные, более короткие, промежутки времени определяется многими факторами: температурой, гидрохимическим режимом, обеспеченностью пищей, состоянием здоровья, особенностями обмена на разных этапах развития, полом, видовой принадлежностью и т. д.
Неравномерность роста зародышей карпа в икринке в период инкубации, т. е. до выклева (в нерестовом пруду хозяйства ТСХА), представлена на рис. 43. Из рисунка видно, что более быстрое нарастание массы охватывает дробление, обрастание желтка, начало формирования органов зародыша, т. е. время усиленного формирования и дифференцировки тканей. После закладки органов следует сильное замедление весового роста. Резкий подъем последнего отрезка кривой показывает быстрое увеличение массы зародыша перед выклевом.
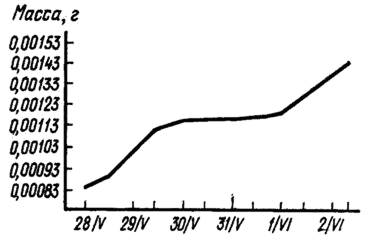
Рис. 43. Периодичность роста эмбриона карпа в период инкубации
Рост молоди в начальный период жизни после выклева также происходит неравномерно. Именно в первые 25—30 дней после выклева выявляются четыре цикла, в которых повышенная скорость роста чередуется с замедленной (табл. 11).
Таблица 11
Периодичность роста молоди карпа в нерестовых прудах (ТСХА)
| Возраст молоди , дни | Длина тела, мм | Масса Р, мг | Кратность увеличения массы | Цикл развития |
| 5,5 | 1,2 | |||
| 2-3 | 5,5 | 1,2 | ||
| (переход на питание) | 6,8 | 1,5 | I | |
| 8,0 | 6,1 | II | ||
| 10,0 | 16,3 | 2,7 | II | |
| 13,5 | 30,0 | III | ||
| 20,0 | 150,0 | III |
I цикл—от вылупления личинок до 4—6-дневного возраста; характеризуется наиболее высокой интенсивностью роста и питания. Биологической особенностью этого цикла является зависимость основных функций организма — питания, дыхания — от желточного мешка, поставляющего организму пищу и кислород.
II цикл—от 4—6-дневного до 6—10-дневного возраста; характеризуется значительной депрессией дыхания и роста. Резорбция желточного мешка вместе с его кровеносными сосудами приводит к понижению функций дыхания и эндогенного питания. Газообмен происходит с помощью личиночных органов. К активному питанию личинки еще не приспособились, они питаются неинтенсивно и в той или иной степени испытывают недостаток пищи. За это время масса тела увеличивается примерно в 2,7 раза.
III цикл — от 10-дневного до 18—20-дневного возраста — новый подъем дыхания и роста. Сформированы жабры, органы пищеварения, передвижения (плавники) и т. д. Увеличение массы за это время — примерно в пять раз.
IV цикл — от 18—20-дневного до 27—30-дневного возраста; заканчивается формирование личинки в малька, появляется чешуя. При обильном питании скорость роста сохраняется высокая, при скудном корме — резко снижается.
Из этих данных следует, что циклы III и IV, характеризующиеся повышенной скоростью роста, являются очень ответственными в раннем развитии рыб. Обильное кормление на этих стадиях обеспечивает хороший рост, а скудное питание приводит к общей депрессии роста. Поэтому нужно пересаживать молодь из нерестовых прудов в выростные во время цикла II, пока рост замедлен, т. е. до 8-дневного возраста, чтобы цикл III проходил уже в условиях обилия пищи. В этом случае будет использована высокая энергия роста организма, способность его к быстрому росту в это время.
У рыб, как у животных с непостоянной температурой тела, наблюдается периодичность, неравномерность роста в течение года. Периоды усиления и замедления роста чередуются в течение года и сезона в соответствии с наиболее или наименее благоприятными для данного вида температурными условиями, богатством или бедностью пищевой базы и т. д. У неполовозрелых карповых, сиговых и многих других рыб характер роста наиболее ясно отражает обеспеченность их пищей и, следовательно, подвержен резким колебаниям. У хищных, например окуневых, рыб зависимость роста от обеспеченности пищевыми организмами выражена слабее. Это связано с тем, что в случаях обеднения пищевой базы они поедают себе подобных (каннибализм).
Очень специфичен рост рыб, которые перестают питаться в период зимовки (большинство объектов разведения в тепловодных хозяйствах) или миграций (проходные рыбы): у них перед зимним голоданием или длительным странствованием в тканях тела накапливается жир при замедленном белковом росте. При этом обычно после зимовки, в начале вегетационного периода, увеличения линейных размеров тела не происходит, а преобладает накопление жира. Этот процесс продолжается до момента достижения определенной упитанности. После этого жиронакопление замедляется или приостанавливается, а ускоряются белковый рост и связанный с ним линейный прирост.
Таким образом, очень важным показателем качества посадочного материала, например карпа, является его упитанность, в какой-то мере отражающая уровень накопления резервных веществ, а следовательно, и подготовленности рыбы к зимнему голоданию и дальнейшему росту во второе лето. Чем упитаннее годовики после зимовки, тем скорее начинается увеличение их линейных размеров после посадки в нагульные пруды.
Рост рыбы в значительной мере отражает качество и количество Пищи, находимой ею в водоеме. Поэтому, как правило, в течение Вегетационного периода увеличиваются различия в величине молоди, полученной одновременно от одних родителей. Например, при выращивании сеголетков карпа разница в размерах выклевывающейся молоди не превышает нескольких процентов, а осенью, т. е. в конце первого лета жизни, самые крупные экземпляры превосходят по массе самые мелкие зачастую в 10—20 раз. Возрастает изменчивость по массе рыб к осени и в нагульных прудах. При этом чем большие колебания размеров наблюдаются при зарыблении прудов весной, тем более разнокачественной оказывается рыба в конце выращивания.
Условия питания определяют быстроту индивидуального роста рыб, а тем самым и характер роста стада в целом. Богатая кормовая база обеспечивает быстрый рост. При этом все рыбы могут потреблять одинаковую пищу, поэтому они растут относительно ровно и индивидуальная изменчивость в стаде невелика.
При недостатке кормовых организмов скорость роста стада замедляется. Даже небольшие различия в исходной величине позволяют рыбам питаться по-разному и различия в росте увеличиваются, становятся все более заметными, а индивидуальная изменчивость рыб в стаде повышается. Таким образом, рост является обобщающим показателем того, насколько условия обитания отвечают потребностям рыбы.
Однако реакция организма на изменение условий жизни неоднозначна. В условиях обильного питания наряду с ускорением роста повышаются, например, выживаемость, особенно молоди, плодовитость, увеличивается численность вида; обеднение пищевой базы, замедление роста рыб сопровождаются уменьшением плодовитости, нередко проявлением каннибализма и в конечном счете уменьшением численности вида.
Неравномерный в течение года рост рыб (сезонные ритмы) приводит к образованию слоев (колец) на чешуе и костях. Эту особенность используют для определения возраста и расчисления темпа роста рыб в предыдущие годы, для чего исследуют чешую, отолиты, отдельные кости скелета — жаберную крышку, лучи плавников, клейтрум, позвонки и т.д. (рис. 44).
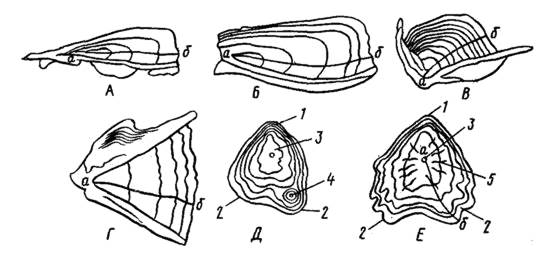
Рис. 44. Кости для определения возраста рыб.
А — клейтрум воблы; Б — клейтрум леща; В — клейтрум осетровых;
Г — крышечная кость окуня; Д, Е — срез луча осетра и севрюги (по Чугуновой,1959):
1 — верхняя часть, 2 — боковые линии, 3 — центр, 4 — добавочный центр,
5 — радиальная бороздка; а—б — линия измерения (в проходящем свете)
У большинства рыб, в том числе карпа, возраст и рост определяют по чешуе. Для приготовления препарата берут несколько чешуек со средней части тела между основанием первого спинного плавника и боковой линией, промывают в слабом растворе нашатырного спирта (1 —10%), обсушивают мягкой тканью и помещают между двумя предметными стеклами. Наилучшие препараты получаются из чешуи, взятой от свежей рыбы, но можно использовать соленую и фиксированную в формалине.
У осетровых, сомов, судака и некоторых других, как правило, используют луч грудного плавника, поперечный срез которого подшлифовывают и приклеивают бальзамом на предметное стекло.
У окуневых, налима и некоторых других рыб удобны для определения плоские кости — жаберные крышки и клейтрум. Косточку очищают, обваривая кипятком, обезжиривают в смеси эфира и бензина в отношении 1:2 и подсушивают. Формалинный материал не используют. Так же подготавливают позвонки, которые раскалывают продольно и рассматривают слом.
У тресковых, камбаловых и судака хорошо определять возраст по отолитам. Отолиты вынимают из свежей рыбы, чаще всего разрезав голову вдоль по середине или поперек в области затылка, разламывают пополам, подшлифовывают и прокаливают. На шлифе хорошо видны годовые кольца.
Для большей достоверности результатов рекомендуют определять возраст рыб параллельно по чешуе и костям. На чешуе и костях видны расположенные друг за другом светлые и темные полосы, или кольца, которые отражают рост рыбы в течение года. Полосы, образующиеся в периоды замедленного роста рыбы, состоят из мелких клеток, плотно прилегающих одна к другой; в падающем свете (освещение сверху) полосы темные, в проходящем (освещение снизу) — светлые. В периоды усиленного роста клетки, образующие полосу, крупнее и лежат свободнее. Кольцо оказывается более широким, матово-светлым в падающем свете и темным — в проходящем.
В умеренных широтах наиболее интенсивное питание и, следовательно, быстрый рост рыб приходится на лето или даже осень, в арктических районах — на зиму, что соответственно отражается на ширине колец. У рыб, обитающих в тропических и экваториальных водах, на костях и чешуе также откладываются сезонные кольца, иногда они соответствуют периодам засухи и тропических ливней.
Широкая и узкая полосы составляют вместе одну годовую зону (рис. 45).
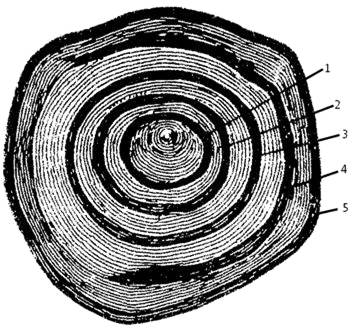
Рис. 45. Чешуя 5-годовалого карпа:
1, 2, 3, 4, 5—годовые кольца
Таким образом, узкие и широкие полосы на чешуе и костях появляются вследствие изменения скорости роста тела. Неправильно называть узкие темные кольца ‛зимними‛: если зимой рыба не питается (карп и др.), то она и не растет и закладки кольца не происходит. Эти кольца соответствуют периодам медленного роста, они могут закладываться весной, осенью и даже летом.
В связи с этим на чешуе и костях могут образовываться не только годовые кольца, но и так называемые дополнительные (добавочные), появляющиеся в результате ослабления или прекращения питания в преднерестовый или нерестовый период, перемены кормовых объектов и т. п. К добавочным кольцам относится прежде всего первое кольцо на чешуе — мальковое, которое возникает при переходе молоди с питания планктоном на питание бентосом. Это кольцо оказывается внутри первой годовой зоны.
У многих рыб, не питающихся во время нереста, этот перерыв в питании отражается на чешуе в виде так называемых ‛нерестовых‛ колец (марок), которые обнаруживаются по разрушенному, как бы размытому краю.
Из-за различия в характере роста рыб до и после наступления половой зрелости у разных возрастных групп время закладки годовых колец сдвигается. Например, у неполовозрелых особей азовского судака и аральского леща, растущих наиболее быстро, годовые кольца закладываются в начале вегетационного периода — еще весной. После наступления половой зрелости определенная доля пищи расходуется на созревание гонад. Поэтому у рыб старших возрастных групп в начале вегетационного периода, когда интенсивно накапливаются резервные вещества, линейный рост часто не происходит. Через определенный промежуток времени жиронакопление замедляется или приостанавливается. Тогда ускоряется линейный рост и начинает закладываться новое широкое кольцо, это происходит уже во второй половине лета.
Таким образом, для правильного ‛чтения‛ чешуи при определении возраста необходимо знать биологию рыб и прежде всего особенности их роста.
Для обозначения разных возрастных групп принята следующая терминология (знак ‛+‛ обозначает прирост следующего года):
| Возрастная группа | Число колец | Обозначение |
| Сеголетки | нет | 0+ |
| Годовики | Одно | |
| Двухлетки | Одно | 1+ |
| Двухгодовики | Два | |
| Трёхлетки | Два | 2+ |
| Трёхгодовики | Три |
И т. д.
Пригодность темных и светлых полос на чешуе и костях рыб для определения возраста открыл еще Левенгук (1684), отметив, что их возникновение связано с неодинаковой быстротой роста. Если знать соотношение роста чешуи (или костей) и роста тела, то можно восстановить линейный рост рыбы в годы, предшествующие поимке (обратное расчисление роста).
Норвежский исследователь Э. Леа на основании допущения, что длина тела рыбы и длина чешуи изменяются в прямой пропорции, т. е. находятся в прямолинейной зависимости ln : l = Vn :V, предложил формулу ln=Vnl /V, где l — длина тела рыбы в момент поимки, V— длина чешуи по средней линии от центра до края, 1n — вычисляемая длина рыбы в возрасте п лет, Vn — расстояние от центра чешуи до годового кольца в возрасте п полных лет.
Установленная зависимость иллюстрируется на рис. 46. Чешую проецируют рисовальным аппаратом на лист бумаги или (в модификации Г. Н. Монастырского) на специальную доску.
Зная длину l рыбы в предыдущие годы, можно вычислить ее еже годные приросты (t): l1, = l1,, t2 = l2—l1 и т. д. Данный метод дает сравнимые результаты.

Рис. 46. Соотношение между скоростью роста рыбы и ее чешуи
(По Никольскому, 1974)
Впоследствии в положения, выдвинутые Э. Леа, были внесены по правки и уточнения; было показано, что между ростом тела и чешуи у некоторых рыб существует не прямая, а логарифмическая зависимость: пропорциональны лишь приращения логарифмов длины тела и длины чешуи. Специальный прибор (Г. Н. Монастырского) позволяет расчислить рост методом логарифмических шкал. Характер образующихся кривых сходен с данными, полученными методом Леа.
При анализе роста рыб пользуются различными показателями в зависимости от поставленной задачи. Обычно вычисляют:
а) весовой или линейный прирост: W1,—W0 (W1—конечная величина, W0 — начальная величина);
б) относительный прирост, или темп роста: (W1,—W0 )/W0 (обо значения те же);
в) относительную скорость роста К (показывает рост в определённый промежуток времени):
к = Wt - Wt /[(Wt + W0 )/2] * t
где W0 величина тела в начале периода,
Wt — величина тела в конце периода,
t — промежуток времени.
Рост рыб взаимосвязан с возрастом: рыбы могут расти практически всю жизнь. Однако, как и у всех живых организмов, с возрастом у них происходят изменения в молекулярной структуре клеток, в соотношении белков, нуклеиновых кислот, липидов, в количестве и качестве ферментов, в способности образовывать антитела. Это существенно влияет на энергетический и пластический обмен всего организма, поэтому его жизнедеятельность и прежде всего очень важные с хозяйственной точки зрения его приспособительные возможности и продуктивность в разном возрасте неодинаковы.
Общие закономерности возраст085;ой изменчивости рыб очень сильно отражаются и на их воспроизводительной способности: во второй половине жизни постепенно снижается плодовитость, а затем, в результате дегенерации гонад, прекращается способность к размножению. Этот процесс сопровождается изменениями качества половых продуктов и свойств получаемого потомства. Поэтому, как и в других отраслях животноводства, в прудовом рыбоводстве большое значение придается возрастному подбору производителей, который в значительной степени определяет формирование качества посадочного материала и затем продуктивность товарной рыбы.
При изучении возрастных изменений основного объекта тепловодного хозяйства (ТСХА) — карпа — было обнаружено, что у производителей разного возраста качество половых продуктов различно. Эти различия касаются количества сухого вещества, белков, жиров, микроэлементов, показателей энергетического обмена. С самого раннего, эмбрионального, периода развития проявляются различия в качестве потомства.
У самцов, созревающих впервые, сперма отличается наименьшим количеством сухого вещества, жира и золы, низким содержанием фосфорных соединений и нуклеиновых кислот, пониженной концентрацией и наименьшим количеством жизнеспособных сперматозоидов в семенной жидкости.
У самцов среднего возраста в сперме отмечено высокое содержание сухого вещества, белков, жиров, аминокислот: цистина, цистеина, триптофана, гистидина; возрастание количества меди и уменьшение цинка. Сперма характеризуется большими запасами нуклеиновых кислот и энергетических веществ (лецитин, холестерин), что свидетельствует о высокой интенсивности обмена. Количество жизнеспособных спермиев в семенной жидкости и общая концентрация спермы наибольшие по сравнению с рыбами других возрастов.
Сперма старых рыб имеет наибольшее количество жира, лецитина, наименьшее — меди, цинка, РНК, пониженное содержание АТФ; понижена также концентрация спермы.
По мере увеличения возраста самок изменяется величина икринок: сначала она возрастает, а затем уменьшается или изменяется незначительно. Меняется и соотношение икринок разных размерных групп: количество крупных икринок сначала увеличивается, а потом уменьшается (табл. 12).
Икра средневозрастных самок содержит наибольшее количество сухого вещества, жиров, протеина и таких аминокислот, как цистин, цистеин, триптофан, гистидин и т. д.; в ней наибольшее количество марганца, меди и цинка.
Характер нереста и основные показатели качества потомства при нересте производителей разного возраста неодинаковы: впервые нерестующие производители нерестуют обычно вяло и недолго (около 3 ч) или, наоборот, очень долго, с промежутками, в общей сложности больше суток. Икра отличается наименьшим оплодотворением (61 — 88,8%), наибольшим отходом в период инкубации, доходящим иногда до 100%, большей продолжительностью инкубации. Выход молоди от одной самки из нерестовых прудов крайне низок и неравномерен (0—90 тыс. шт.). Замедленный рост и большие отходы характеризует их потомство и при дальнейшем выращивании, в первое и второе лето жизни.
У производителей, нерестующих второй раз, нерест длится дольше, он более активный. Оплодотворённость икры повышается (93,3%). Отход икры ниже, чем при первом нересте, но все же высок (14,3%). Выход молоди от одной самки из нерестовых прудов 31 —135 тыс. шт.
Производители среднего возраста нерестуют бурно, в течение 5—8 ч. Икра оказывается наиболее оплодотворенной (91—98%), эмбриональное развитие потомства проходит быстро.
У производителей среднего возраста отмечен наименьший отход икры, в ней больше жиров и белка, выклев молоди и переход ее на внешнее питание проходит быстрее. От каждой самки из нерестовых прудов получают наибольшее количество молоди (свыше 150 тыс. шт.). В постэмбриональный период эта молодь сохраняет преимущества по основным рыбоводным и биохимическим показателям (отходы, рост, накопление жиров, прирост белка и т. д.).
Так относительное увеличение массы зародышей при инкубации икры в зависимости от возраста производителей выглядит следующим образом:
| Возраст производителей, лет1 | Относительное увеличение массы (Р2—Р1)/Р21 | Возраст производителей, лет1 | Относительное увеличение массы (Р2—Р1)/Р21 |
| 0,53 | 0,52 | ||
| 0,28 | 0,40 | ||
| 0,83 | 0,30 |
1- Икру производителей возраста 6 и 10, 7 и 11, 8 и 12 лет инкубировали одновременно (попарно).
2- P1, и Р2 — начальная и конечная массы зародыша.
Стареющие производители нерестуют менее бурно, долго, с промежутками. Эмбриональное развитие молоди длится дольше, отходы икры повышаются (19—34%), весовой рост ее во время инкубации и желточного питания идет медленнее. Выклев и переход на активное питание растягивается. Выход молоди от одной самки из нерестовых прудов невысок (67—81 тыс. шт). При дальнейшем выращивании молодь характеризуется повышенным отходом.
При дальнейшем старении карпов способность их к размножению затухает: некоторые пары перестают нерестовать в возрасте 13—15 лет. Повышаются отходы икры, молодь отстает в росте.
По основным рыбоводным показателям — выживаемости, росту, зимостойкости — лучшие сеголетки и годовики получаются от производителей среднего возраста, худшие — от впервые нерестующих, поэтому рыбопродуктивность при выращивании потомства от производителей разного возраста различна.
Дата добавления: 2015-06-22; просмотров: 2270;