Размножение и развитие рыб
Развитие организма представляет собой совокупность количественных и качественных изменений в результате взаимодействия организма со средой. В индивидуальном развитии рыб можно выделить ряд крупных отрезков – периодов, каждый из которых характеризуется общими для разных видов свойствами.
I. Эмбриональный период – oт момента оплодотворения яйца до перехода молоди на внешнее питание. Эмбрион питается за счёт желтка – запаса пищи, полученного от материнского организма. Этот период подразделяется на два подпериода:
1) подпериод икринки, или собственно эмбриона, когда развитие происходит в оболочке;
2) подпериод свободного эмбриона (предличинки), когда развитие идет вне оболочки.
II. Личиночный период начинается с момента перехода на питание внешней пищей; внешний облик и внутреннее строение ещё не приняли формы взрослого организма. У личинок имеются специфические личиночные органы, которые в дальнейшем пропадают.
III. Мальковый период – внешний облик близок к облику взрослого организма. Исчезают личиночные органы, появляются характерные для взрослых органы и функции. Половые органы почти неразвиты. Энергетические ресурсы расходуются главным образом нарост. Вторичнополовые признаки обычно отсутствуют.
IV. Период полувзрослого (неполовозрелого) организма: начинается более или менее бы 89;трое развитие половых желез и вторично-половых признаков, но организм ещё не способен к размножению.
V. Период взрослого (половозрелого организма – состояние, при котором в определённый период года организм способен воспроизводить себе подобных; вторичнополовые признаки, если они свойственны данному виду, имеются. Энергия тратится преимущественно. На развитие половой системы и создание запасов для поддержания жизнедеятельности во время миграций, зимовок, размножения.
VI. Период старости – половая функция затухает; рост в длину прекращается или крайне замедляется.
В пределах подпериода или периода выделяют этапы. Теория этапности развития рыб разработана отечественными учеными школы академика А. Н. Северцова С. Г. Крыжановским и В. В. Васнецовым. На каждом этапе организм характеризуется специфическими приспособлениями к среде, т. е. определёнными особенностями строения, дыхания, питания, роста. На протяжении этапа организм растет, но значительных изменений в его строении и отношениях со средой не происходит. При этом вырабатываются свойства, обеспечивающие переход к следующему этапу. Стадия обозначает любой данный момент развития.
Размножение – важнейший жизненный процесс, обеспечивающий существование вида. В органическом мире размножение может происходить двумя способами – бесполым и половым.
Рыбам свойственно половое размножение, хотя у многих видов сельдей, осетровых, лососевых, карповых и некоторых других зрелые половые клетки, попав в воду, начинают развиваться партеногенетически, т. е. без оплодотворения. При этом, как правило, развитие доходит только до стадии дробления и лишь в исключительных случаях были получены жизнеспособные личинки, прожившие до полного рассасывания желточного мешка (салака, сахалинская сельдь, налим, чебачок, окунь).
Некоторым видам рыб (серебряный карась Carassius auratusgibelio, молинизия Molinisia formosa) свойствен совершенно необычный способ размножения – гиногенез. У этих видов во многих районах ареала популяции состоят только из самок (самцы отсутствуют или единичны и неполноценны в половом отношении). В таких однополых популяциях нерест самок проходит при участии самцов других видов. При этом виде размножения проникновение спермия в яйцеклетку является необходимым условием развития. Однако слияния ядер спермия и яйцеклетки не происходит и ядро яйцеклетки становится ядром зиготы (ядро спермия генетически инактивируется). В результате в потомстве появляются только самки без внешних признаков тех самцов, которые принимали участие в нересте. Цитогенетической основой этого процесса является триплоидия самок из однополых популяций (при специфическом ходе делений созревания).
Размножение и развитие рыб отличаются рядом специфических особенностей, обусловленных водным образом жизни.
У большинства рыб осеменение наружное. В отличие от наземных животных зрелые половые клетки рыб выводятся в воду, здесь происходит оплодотворение икры и дальнейшее ее развитие. Осеменение, оплодотворение и инкубация икры в воде, вне материнского организма, влечет за собой большую гибель потомства на ранних стадиях развития. Для обеспечения сохранения вида в процессе эволюции у рыб выработалась или большая плодовитость, или забота о потомстве.
Плододовитость рыб много выше, чем у наземных позвоночных. Это приспособительное свойство вида к условиям существования. Количество икры, откладываемой разными видами, очень сильно варьирует – от нескольких штук у полярной акулы до 200 млн. у морской щуки и 300 млн. у луны-рыбы. Наиболее плодовиты рыбы, откладывающие плавающую пелагическую икру; затем следуют рыбы, икра которых развивается приклеенной к растениям. У рыб, прячущих или охраняющих свою икру, плодовитость невелика.
Наблюдается обратная зависимость между индивидуальной плодовитостью и размерами икринок: у рыб с крупной икрой она ниже, с мелкой – выше (у кеты диаметр икринок 7–8 мм, плодовитость2–4 тыс. шт. , у трески диаметр икринок 1,1 –1,7 мм, плодовитость до 10 млн. шт. ).
Сильнейшее влияние на плодовитость оказывает обеспеченность рыб пищей. У рыб одного размера плодовитость значительно выше в благоприятных условиях питания – в кормные годы, в разреженных популяциях. Кроме того, у одного и того же вида плодовитость зависит от размера и возраста рыбы. У одной и той же особи плодовитость –при прочих равных условиях – по мере роста сначала увеличивается, затем к старости уменьшается, несмотря на продолжающийся абсолютный рост.
Различают плодовитость индивидуальную, относительную и рабочую. Индивидуальная, или абсолютная, или общая, плодовитость –это общее количество икры, выметываемое самкой за один нерестовый период при благоприятных условиях. Например, у 6-летнегокарпа она составляет в среднем около 900 тыс. шт. Относительная плодовитость – количество икры, приходящееся на единицу массы тела самки; у карпа 180 тыс. шт/кг массы тела; этот показатель особенно удобен для сравнения, он четко показывает, как изменяется количество икры с ростом рыбы: до определённого возраста оно возрастает, потом снижается. Рабочая плодовитость – количество икры, получаемое от одной самки при проведении искусственного осеменения в рыбоводной практике. Для определения плодовитости берут пробу икры в стадии ее наиб;ольшего развития, т. е. незадолго до нереста.
Наступление половой зрелости у разных видов происходит в разном возрасте, причем во многих случаях самцы созревают на год раньше самок. Наиболее скороспелые промысловые рыбы, с коротким жизненным циклом (некоторые бычки, каспийская килька, хамса, снеток), созревают в годовалом возрасте. Рыбы с продолжительным жизненным циклом, например осетровые, становятся половозрелыми в 7–8 (севрюга), 12–13 (осетр) и даже 18–20 лет (белуга и калуга).
У рыб одного и того же вида половозрелость может наступать в разном возрасте, в зависимости прежде всего от температуры выращивания и наличия пищи. Карп в средней полосе созревает в возрасте 4–5 лет, в южных районах – в 2 года и затем нерестует1 раз в год, в тропиках – на Кубе – в возрасте 6–9 месяцев и нерестует с полугодовым интервалом. У рыб одного вида ритм полового созревания в тропиках иной, чем в средней полосе (рис. 33). Обычно время наступления половой зрелости связано с достижением особью определённых размеров. Чем медленнее рыба растет, тем позднее созревает. Соотношение полов у разных видов изменчиво, но у большинства близко к 1: 1, кроме тех, у которых наблюдается гиногенез.

Рис. 33. Ритм половых циклов рыб (карпа) в разных широтах.
А – в тропиках; Б– в умеренной зоне (по Привезенцеву, 1981):
1 – I стадия, 2 – II, 3 – III, 4 – IV, 5 – V, 6 – VI стадия
Вторичных половых признаков у большинства рыб нет, поэтому у них самки и самцы внешне не различаются. Однако половой диморфизм проявляется в различной величине самок и самцов: среди одновозрастных рыб самки бывают крупнее самцов, за исключением некоторых видов – мойвы, нерки, канального сома. Однако у многих рыб половой диморфизм становится Заметным в преднерестовый период, при созревании, когда появляется так называемый брачный наряд – в подавляющем большинстве случаев у самцов. У карповых и сигов на голове и теле образуется жемчужная сыпь, у хариусов краснеют плавники, у гольцов на теле появляются яркие пятна, у лососей изменяются челюсти и возникает горб (нерка, горбуша), появляются лиловые пятна вдоль тела (кета) и т. д. После нереста брачный наряд пропадает, однако, например, у дальневосточных лососей, угрей, сельди-черноспинки изменения в организме настолько сильны и необратимы, что после первого нереста рыбы погибают. У некоторых видов нашей фауны вторичные половые признаки выражены достаточно четко.
Они проявляются в большинстве случаев в величине плавников: у некоторых сомов, пескаря Gobio gobio, байкальской желтокрылки Cottocomephorus grewingkii, грудные плавники у самцов больше, чем у самок; у самцов линей Tinca tinca брюшные плавники больше и вторые лучи их толще; у самцов хариуса Thymallus thymallus и четырехрогого подкаменщика Myoxocephalus quadricornis длиннее спинные плавники. У полярных камбал рода Liopsetta самцы имеют ктеноидную, а самки – циклоидную чешую. У некоторых глубоководных рыб (Ceratiidae) мелкие самцы прирастают к телу самки.
Очень своеобразны преднерестовые изменения у горчаков Rhodeinae, откладывающих икру в мантийную полость двустворчатых моллюсков: у самцов окраска становится очень яркой, а у самок вырастает длинный яйцеклад.
По срокам икрометания рыб нашей фауны разделяют на:
а) весенне-нерестующих (сельди, радужная форель, щука, окунь, плотва, орфа);
б) летне-нерестующих (сазан, карп, линь, краснопёрка);
в) осенне-зимне-нерестующих (многие лососи, сиги, налим, навага).
Это деление в известной мере условно–один и тот же вид в разных районах нерестует в разное время: карп нерестует в средней полосе в мае–июне, на островах Ява и Куба – круглый год.
Время нереста сильно варьирует в течение суток: лососи, налим, хамса обычно вымётывают икру ночью, анчоус – вечером, карп нерестует чаще всегl6; на зорях.
Изменения половых желез рыб в течение года (годичные половые циклы) проходят по одной схеме. Однако у разных видов имеются особенности в созревании и прежде всего в продолжительности разных стадий зрелости.
По продолжительности периода икрометания выделяют две группы рыб: с единовременным и порционным нерестом. У рыб единовременного икрометания икра откладывается сразу, единовременно: в короткий срок (одно утро) нерестуют вобла, окунь. Многие тропические рыбки выметывают икру в течение часа. Вся икра таких рыб, предназначенная к вымету в данный сезон, созревает сразу и полност100;ю выметывается.
Другие рыбы откладывают икру в несколько приемов, отдельными порциями, с промежутками в 7–10 дней. Типичный представитель – каспийские сельди. У них в гонадах икра созревает и выметывается последовательными порциями. В результате увеличивается индивидуальная плодовитость: при порционном икрометании за сезон самка выметывает в 2–3 раза больше икры, чем при единовременном.
Порционность икрометания характерна главным образом для рыб тропиков и субтропиков, в умеренных широтах их меньше, в Арктике – почти нет.
Существуют рыбы, которые хотя и не имеют резко выраженного порционного икрометания, но нерестовый период их (одной особи) растягивается на несколько дней, т. е. икра также выметывается в несколько приемов (лещ, иногда карп). Некоторые рыбы в южной части своего ареала нерестуют порционно, в северной – единовременно (лещ, карп).
Порционное икрометание способствует увеличению плодовитости рыб и обеспеченности потомства пищей, а также лучшей выживаемости молоди в неблагоприятных условиях обитания. Например, в водоемах с колеблющимся уровнем значительно больше видов рыб с порционн 99;м нерестом.
Выметанные икринки у подавляющего большинства рыб шаровидны, но есть и овальные (хамса), сигаровидные (бычки, ротан) и даже каплевидные и цилиндрические (некоторые бычки). Окраска икринок у большинства видов желтоватая, оранжевая разных оттенков, у осетровых – черная, у бычков – зеленая. Желтоватый и оранжевый цвет обусловлен присутствием каротиноидов. Размеры икринок сильно варьируют: у некоторых сельдей, камбал икринки имеют менее 1 мм в диаметре, у акул – до 8–9 см и выше, причем они увеличиваются по мере продвижения вида к северу и на глубины.
Величина икринок, мм
Плащеносная акула Chlamydoselachus anguineus 90 – 97
Ручьевая форель Salmo trutta morpha fario 4,0 – 6,5
Черноморский шпрот Sprattus sprattus 0,9 – 1,15
Сельдь-черноспинка Caspialosa kessleri 2,87 – 3,93
Карп Cyprinus carpio 0,9 – 1,5
Линь Tinca tinca 1,0 – 1,2
Чехонь Pelecus cultratus 3,8 – 5,9
Речная камбала Pleuronectes flesus 0,78 – 1,3
Окунь Perca fluviatilis 2,0 – 2,5
Судак Lucioperca lucioperca 1,2 – 1,4
Щука Esox lucius 2,5 – 3,0
Икринки, выметанные и развивающиеся в разных экологических условиях, обладают рядом особенностей, которые способствуют их приспособленности к среде, (рис. 34). В толще воды развиваются плавающие, или пелагические, икринки, на дне или на субстрате –донные, или демерсальные.
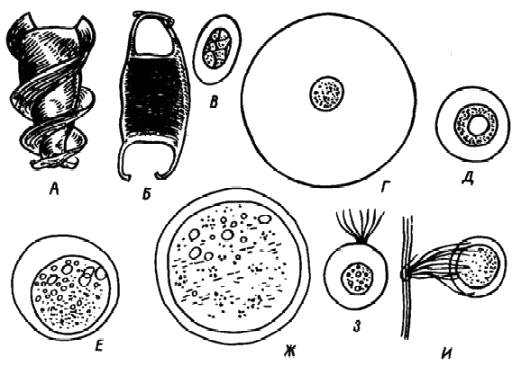
Рис. 34. Икринки, развивающиеся в разных экологических условиях (масштаб не соблюдён).
А, Б – акул и скатов (по Никольскому, 1971); пелагические: В–хамсы, Г – чехони, Д – змееголова;
донные: Е – белорыбицы, Ж – форели; прикрепленные: З – сайры, И – атерины
У пелагических икринок, развивающихся в толще воды, увеличение плавучести обеспечивается рядом приспособлений. К ним относятся: оводнение желтка (в морских пелагических икринках содержание воды доходит до 85–97%, благодаря чему они легче морской воды, тогда как в донных – до 60–76%), увеличение перивителлинового пространства за счет наличия в желтке жировых капель (многие сельди, камбалы) или образование выростов, облегчающих удерживание икринки в толще воды (сайра и др. ).
У чехони, дальневосточных растительноядных рыб, проходных сельдей икринки полупелагические; они развиваются в толще воды, на течении, в реке, но в стоячей воде тонут.
Икринки, откладываемые на субстрат (вегетирующие или отмершие растения, камни, коряги и т. д. ), часто обладают клейкими оболочками (осетровые, атлантическая и тихоокеанская сельди, карп, карась, рыбец и т. д. ) или снабжены нитевидными или крючковидными отростками, которыми они прикрепляются к субстрату. Икринки часто откладываются компактно, и кладки имеют характерную форму. Например, у окуня икринки окружены вязким студенистым веществом, кладки имеют вид длинных (2–3 м) лент (рис. 35). Однако они могут и не прикрепляться к субстрату (лососевые, налим). Донные икринки свойственны подавляющему большинству пресноводных рыб или морским, нерестующим в прибрежной зоне. Количество желтка и плазмы в икринках разных видов рыб не одинаково. По их соотношению яйца костистых рыб делят на олигоплазматические (содержащие мало плазмы и много желтка) и полиплазматические (богатые плазмой и бедные желтком).
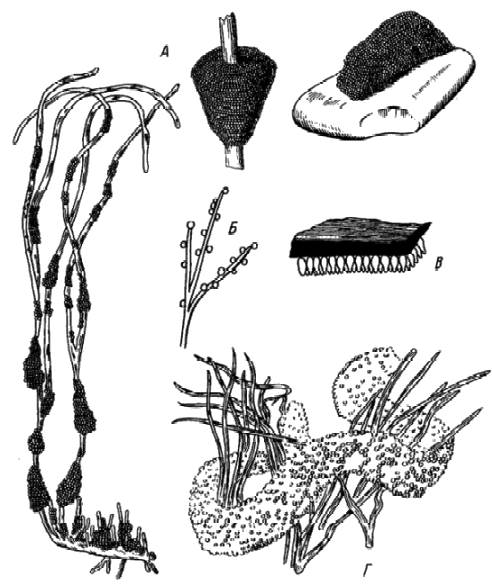
Рис. 35. Кладки икры тихоокеанской сельди (А), карпа (В), ротана (В), окуня (Г)
Резервный материал для питания зародыша – желток овоцита –состоит преимущественно из белков, основная масса которых представлена липофосфопротеидами (ихтулин) и небольшим количеством альбумина, и липидов (главным образом фосфатидов, прежде всего лецитина, а также холестерина); имеется небольшое количество полисахаридов и нейтральных жиров.
У многих рыб цитоплазма овоцита содержит жировые капли, состоящие преимущественно из нейтральных жиров – глицеридов. Яйца рыб характеризуются большим количеством воды. Сильно колеблется в них содержание белков (от 12–14 до 29–30% от сырой массы) и жиров (от 1–2 до 22% сырой массы). При этом количество белков преобладает над; количеством жиров (отношение белки/жиры, например у пеляди составляет 1,17, у форели – 3,25, у сазана – 4,15, а у щуки и судака – 21,19–21,66).
Различна и калорийность икры, например у осетра и лосося 25522–25941 Дж/г, у барабули 16 318 Дж/г сухого вещества. Содержание углеводов в яйцах рыб незначительно: глыбки гликогена обнаружены (цитохимически) у осетровых, лосося, пеляди, карпа.
Таким образом, главным источником энергии при развитии зародыша являются белки, за счет которых покрывается до 70% расходуемой энергии. Жиры, в отличие от жиров яиц птиц, расходуются в меньшей степени. При этом в пелагических икринках запас энергетических ве097;еств меньше, в донных – больше.
Зрелый сперматозоид представляет собой клетку с малым количеством плазмы. В нем различают головку, среднюю часть и хвост (рис. 36). Форма головки различна: в виде шара, яйца, желудя (у большинства костистых рыб), палочки (у осетровых и некоторых костистых), копья (у двоякодышащих), цилиндра (у акуловых, кистепёрых). В головке помещается ядро. Впереди ядра у акулообразных, осетровых и некоторых других рыб располагается акросома; у костистых акросомы нет. Ядерная часть головки сперматозоида состоит преимущественно из дезоксирибонуклеопротеида (нейтральная соль ДНК с основным белком – протамином) и небольшого количества РНК. Концентрация ДНК в головке (ядре) составляет 38,1% (карп), 48,4% (семга) и отражает количество ДНК в гаплоидном наборе хромосом. Протамины состоят из 6–8 аминокислот, среди которых преобладает аргинин. В средней части сперматозоидов обнаружены митохондрии, играющие основную роль в снабжении клетки энергией. В хвостовой части выявлены белки, лецитин, жиры и холестерин. Сперматозоиды большинства костистых рыб имеют общую длину 40–60 мкм (головка 2–3мкм).

Рис. 36. Сперматозоиды рыб.
А – костистых; Б – осетра (по Гинзбургу, 1968):
1 – карася, 2 – щуки, 3 – бычка Gobius niger,
4 – подкаменщика Cottidae (вид головки с уплощённой стороны и сбоку);
a – акросома, г – головка вместе со средней частью,
гч – главная часть, кч – концевая часть хвоста
Сперма, выделяемая самцом, состоит из сперматозоидов, погруженных в спермиальную жидкость, сходную по составу с физиологическим раствором. В момент выхода из организма сперматозоиды ещё неподвижны, обмен их понижен.
У одного и того же самца сперматозоиды качественно неодинаковы. Прежде всего они различаются по величине: в эякуляте при помощи центрифугирования можно выделить три группы сперматозоидов – мелких (легких), крупных (тяжелых), промежуточных (средних). Они различаются и по биологическим свойствам, в частности по характеру гамет: среди крупных сперматозоидов в большом количестве встречаются X-гаметы, среди мелких – Y-гаметы. Вследствие этого из икры, оплодотворенной крупными сперматозоидами, рождаются преимущественно самки, а мелкими – самцы.
Эти данные используют для разработки направленного формирования пола у рыб, которое имеет важное значение в рыбоводстве. Количество спермы, которое единовременно выделяет самец при нересте, и концентрация эякулятов различны у разных видов (табл. 6). Оно зависит от комплекса внутренних и внешних факторов: размеров, возраста и состояния самцов, условий нереста – течений, температуры, соотношения самок и самцов на нерестилищах и т. д.
В спермиальной жидкости сперматозоиды неподвижны. При с086;прикосновении с водой их обмен резко возрастает, поглощение кислорода увеличивается больше чем вдвое, содержание АТФ – больше чем втрое; сперматозоиды активируются и начинают бурно двигаться со скоростью, наблюдаемой у сперматозоидов млекопитающих (164–330 мкм/с). Встретив икринки, они проникают в них через микропиле, после чего происходит оплодотворение. Однако в воде сперматозоиды недолговечны. Энергетические ресурсы их иссякают, первоначальное поступательное движение замедляется, становится колебательным, затем прекращается и они погибают. Продолжительность подвижности сперматозоидов колеблется в зависимости от солености среды, в которой проходит нерест, и температуры; в соленой воде она значительно дольше: до нескольких суток у тихоокеанской сельди Clupea harengus pallasi, в пресной воде у большинства рыб – карповых, лососевых, окуневых – не больше 1–3 мин.
Таблица 6
Характеристика спермы разных видов рыб
(по Казакову, 1978, с сокращениями)
| Вид рыбы | Единовремен ный объём спермы, см3 | Концентрация сперматозоидов, млн. /мм | Продолжительность активности сперматозоидов, с 1 2 |
| Радужная форель Пелядь Карп Белый амур | 1,0-23,0 0,2-3,2 2,9-12,5 20-30 | 20,4 7,6 16,5 33,1 | 60-105 10-56 - 27-65 - 70-87 35 15-53 |
Общая
Дата добавления: 2015-06-22; просмотров: 4794;