Механизмы мембранного транспорта
Липидные бислои в значительной степени непроницаемы для подавляющего большинства веществ, и поэтому перенос через липидную фазу требует значительных энергетических затрат.
Различают активный транспорт и пассивный транспорт (диффузию).
Пассивный транспорт – это перенос молекул по концентрационному или электрохимическому градиенту, т.е. он определяется только разностью концентраций переносимого вещества на противоположных сторонах мембраны или направлением электрического поля и осуществляется без затраты энергии АТФ. Возможны два типа диффузии: простая и облегченная (рис. 4.5).
1. Простая диффузия. Происходит без участия мембранного белка. Скорость простой диффузии хорошо описывается обычными законами диффузии для веществ, растворимых в липидном бислое. Скорость движения молекулы определяется концентрационным градиентом и растворимостью молекулы в липидах. Механизм диффузии водорастворимых веществ менее изучен. Перенос вещества через липидный бислой, например, таких соединений, как этанол, возможен через временные поры в мембране, образованные разрывами в липидном слое при движении мембранных липидов. В мембранах также существуют каналы, образованные белками, через которые могут двигаться молекулы. По механизму простой диффузии осуществляется трансмембранный перенос газов (например, О2 и СО2), воды, некоторых простых органических ионов и ряда низкомолекулярных жирорастворимых соединений. Следует помнить, что простая диффузия осуществляется неизбирательно и отличается низкой скоростью.
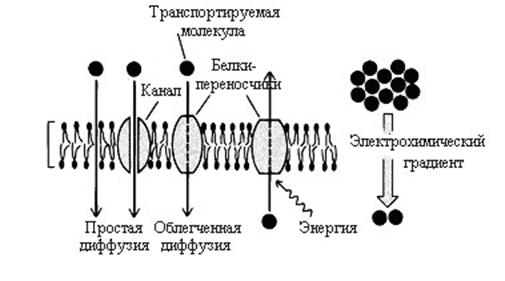
Рис. 4.5. Механизмы пассивного транспорта.
2. Облегченная диффузия – движение молекул по градиенту концентрации с использованием специфических мембранных белков-переносчиков. Следовательно, облегченная диффузия – это диффузионный процесс, сопряженный с химической реакцией взаимодействия транспортируемого вещества с белком-переносчиком. Этот процесс специфичен и протекает с более высокой скоростью, чем простая диффузия. Скорость переноса определяется концентрационным градиентом через мембрану и количеством молекул переносчика.
Известны два типа мембранных транспортных белков: белки-переносчики, называемые транслоказами или пермеазами, и белки каналообразующие.
3. Активный транспорт – это транспорт веществ против градиента концентрации, с использованием переносчика и затратой энергии. Источником энергии может быть АТФ, градиент концентрации или солнечная энергия. Различают первичный активный транспорт и вторичный активный транспорт.
В первичном активном транспорте используется энергия гидролиза АТФ. Известны три основных типа первичного активного транспорта:
1) Натрий-калиевый насос – Na/K+- аденозинтрифосфатаза (Na/K+-АТФ-аза) переносит ионы натрия из клетки, а калия – в клетку.
2) Кальциевый насос – Ca2+-АТФ-аза, который транспортирует Са2+ из клетки или цитозоля в саркоплазматический ретикулум.
3) Н+-АТФ-аза – протонный насос, функционирующий в сопрягающих мембранах, в том числе в митохондриальной мембране.
Для активного транспорта, как и для облегченной диффузии, характерны высокая специфичность, эффект насыщения транспортных белков транспортируемыми молекулами, а также действие ингибиторов.
В качестве примера первичного активного транспорта можно привести транспорт, осуществляемый Na+,K+-АТФ-азой, как одной из наиболее важных и широко распространенных активных транспортных систем в плазматической мембране животных- клеток.
Клетка содержит низкую концентрацию Na+ (в 10 раз ниже) и высокую концентрацию К+ (в 30 раз выше), чем в окружающей среде. Na+,K+-АТФ-аза была открыта в 1957 г. Й.Скоу во фракции плазматических мембран нервов краба, впоследствии она была обнаружена во всех исследованных клетках животных, особенно велико ее содержание в органах, осуществляющих солевой обмен (почки) или выполняющих электрическую работу (мозг, нервы).
Na+,K+-АТФ-азой является интегральным мембранным белков (м.м. 250 000). Состоит из 2-х α и 2-х β субъединиц. α-Субъединица пронизывает мембрану насквозь и имеет центры связывания для АТФ и Na+ на цитоплазматической стороне мембраны и центры для связывания с К+ на внешней стороне. β-субъединицы содержат углеводные группы, расположенные на наружной стороне плазматической мембраны. Она способствует правильной ориентации ферментов в липидном бислое. Перенос ионов происходит за счет изменения конформации фермента при его фосфорилировании-дефосфорилировании за счет АТФ.
По общепринятому представлению, механизм действия АТФ-азы включает несколько стадий.
1. Присоединение 3Na+ вызывает активацию АТФ-азы, происходит гидролиз АТФ и фосфорилирование фермента.
2. Фосфорилирование фермента вызывает изменение конформации и открытие канала снаружи. АТФ-аза теряет сродство к ионам натрия и 3Na+ выводятся через канал на наружную сторону.
3. Два иона К+ присоединяется к ионсвязывающим центрам фосфорилированного белка.
4. Происходит (возможно, самопроизвольный) гидролиз фосфоэфирной связи и дефосфорилирование фермента, что вызывает его переход в исходную конформацию (открытие канала внутри).
5. Происходит снижение сродства к ионам К+ и выход их в цитозоль и вновь присоединяется АТФ и 3 Na+.
Неравнозначный перенос заряженных ионов (частиц) через мембрану вызывает ее поляризацию: появление «+» снаружи и «-» изнутри. Создаваемый градиент используется для вторичного активного транспорта, например, глюкозы в клетки. Переносчик глюкозы обеспечивает транспорт глюкозы в клетку кишечника за счет входа в клетку ионов Na+ под действием электрохимического градиента (концентрация Na+ высокая в просвете кишечника и низкая в цитозоле клеток). Глюкоза из клетки переходит во внеклеточную жидкость по механизму облегченной диффузии. Nа+,К+-АТФаза поддерживает эту концентрацию Na+ за счет его откачки в межклеточное пространство в обмен на ионы К+ с затратой АТФ.
Дата добавления: 2015-06-12; просмотров: 3189;