Структура мембран
Все биологические мембраны имеют общие признаки строения.
1. Мембраны являются сложными структурами, построенными из липидов, белков и углеводов.
2. Соотношение белков и липидов в мембранах варьирует от 1:4 до 4:1 и зависит от типа клеток и органелл.
3. Мембраны являются асимметричной структурой с наружной и внутренней поверхностями.
4. Мембрана стабилизируется нековалентными связями и является термодинамически стабильной и метаболически активной.
5. Специфические белки встроены в мембраны и выполняют специфические функции рецепции управляющих сигналов, межклеточного взаимодействия, транспорта веществ и пр.
Основными липидами мембран являются фосфолипиды, гликолипиды и холестерин.
1. В мембранах присутствуют 2 основных класса фосфолипидов:
1) Глицерофосфоглицериды являются основным компонентом большинства мембран и состоят из спирта глицерина, 2-х остатков жирных кислот и фосфорилированного спирта. Жирных кислоты содержат четное число атомов углерода (чаще 16 и 18). Жирные кислоты неразветвленные и могут быть насыщенными (чаще в 1 положении) и ненасыщенными (чаще во 2 положении).
2) Сфингомиелины содержат спирт сфингозин. Жирная кислота присоединяется амидной связью к аминогруппе сфингозина. Первичная гидроксильная группа сфингозина эстерифицируется фосфорилхолином. Сфингомиелины находятся преимущественно в миелиновых оболочках.
2. Гликолипиды являются сахаросодержащими липидами и делятся на ганглиозиды и цероброзиды. Цереброзиды вместо фосфорилированного спирта содержат остаток гексозы (глюкозу или галактозу). Ганглиозиды содержат цепь из 3 и более остатков углеводов (например, сиаловые кислоты), которые присоединяются к первичной спиртовой группе сфингозина.
3. Стероиды. Основным стероидов в мембранах является холестерол (рис. 4.1), который находится преимущественно в плазматической мембране клеток.
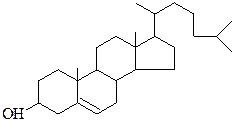
Рис.4.1. Холестерол
Холестерол в основном встречается в наружном слое плазматической мембраны. Холестерол встроен между молекулами фосфолипидов так, что его гидроксильная группа обращена к водной поверхности, в гидрофобное кольцо находится в толщине липидного слоя.
Мембрана является амфипатической структурой.
1.Все молекулы липидов, входящих в состав мембран, имеют гидрофильную и гидрофобную области и поэтому являются амфипатическими.
2.Амфипатические липиды мембран имеют полярную головку и неполярные гидрофобные хвосты (рис. 4.2).
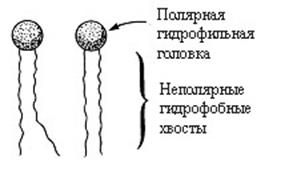
Рис. 4.2. Структура липидов мембран.
3.Насыщенные жирные кислоты имеют прямые хвосты, ненасыщенные жирные кислоты имеют цис-конформацию и изогнутые хвосты, что делает мембрану менее жесткой и более текучей.
4. Липиды формируют бислой, в котором гидрофобная область фосфолипидов защищена от воды, а гидрофильная область обращена к воде (рис. 4.3).

Рис. 4.3. Строение липидного бислоя.
По расположению белков в мембране, способу их ассоциации с липидным бислоем их можно разделить на: поверхностные (периферические) мембранные белки, связанные с гидрофильной поверхностью липидного бислоя; интегральные мембранные белки, погруженные в гидрофобную область бислоя.
1. Периферическиебелки связаны с гидрофильной поверхностью бислоя своими полярными радикалами с образованием нековалентных (ионных, водородных) связей. Многие периферические белки взаимодействуют нековалентными связями с гидрофильной областью интегральных белков. Периферические белки могут быть выделены без разрушения мембраны концентрированными солевыми растворами или изменением рН. Многие рецепторы для гормонов являются периферическими белками.
2. Интегральные белкипогружены в мембрану на определенную глубину или пронизывают мембрану насквозь (прошивающие). Такие белки могут быть экстрагированы только при разрушении мембраны органическими растворителями или детергентами.
Трансмембранные белки могут в виде нескольких α-спиралей или одиночной α-спирали пересекать мембрану. При этом большая (гидрофильная) часть такого белка экспонирована в воду. Некоторые белки могут проникать только на расстояние одного монослоя мембраны.
Интегральные белки, подобно липидам, обладают амфипатическими свойствами у них есть гидрофобные области, взаимодействующие с гидрофобными радикалами липидных молекул внутри бислоя, и гидрофильные, обращенные с обеих сторон мембраны к воде.
Если основные структурные особенности биологических мембран определяются свойствами их липидного бислоя, то специфические функции мембран – белками.
1. На основании роли белков в мембране их можно разделить на две группы: структурныеидинамические белки. Структурные белки поддерживают структуру всей мембраны. Это, как правило, периферические белки, выступающие в роли «молекулярного бандажа». Динамические белки непосредственно участвуют в процессах, происходящих на мембране. Выделяют три класса таких белков:
1) транспортные – участвующие в трансмембранном переносе веществ;
2) каталитические – это ферменты, интегрированные в мембрану и катализирующие происходящие там реакции;
3) рецепторные – это мембранные рецепторы, специфически связывающие такие соединения, как гормоны, нейромедиаторы, токсины, на наружной стороне мембраны, что служит сигналом для изменения метаболических процессов в мембране или внутри клетки.
2.Углеводы мембран. В составе мембран углеводы находятся только в соединении с белками (гликопротеины и протеогликаны) и липидами (гликолипиды). В мембране гликозилировано около 10% всех белков и 5-25% липидов. Углеводные цепи белков колеблются по составу от 2-х членных структур до разветвленных 18-членных полисахаридов.
Функции углеводов:
1) определяют межклеточное взаимодействие;
2) участвуют в системе иммунитета (антигенные детерминанты групп крови);
3) входят в состав рецепторов.
Выделяют основные характеристики мембран:
1.Текучесть (жидкостность) мембраны.Текучесть мембраны характеризуется способностью компонентов мембраны к движению. Текучесть мембраны определяется липидным составом, процентом полиненасыщенных жирных кислот и холестерина. Особое влияние на текучесть мембраны оказывает четырехчленное кольцо холестерола, погруженное в липидный бислой. У эукариотических клеток при температуре 37°С холестерол ограничивает текучесть мембраны, а при более низких температурах он, наоборот, способствует поддержанию их текучести, препятствуя слипанию углеводородных цепей. Текучесть влияет на функцию мембраны. При повышении текучести увеличивается проницаемость для воды и других гидрофильных молекул.
2. Избирательная проницаемость.Это свойство обеспечивает регуляцию транспорта в клетку необходимых молекул, а также удаления из клетки продуктов метаболизма, т.е. активный обмен клетки и ее органелл с окружающей средой. Избирательный транспорт необходим также для поддержания трансмембранного градиента ионов, служит основой всех биоэнергетических механизмов, определяет эффективность процессов рецепции, передачи нервного возбуждения и т.п.
3. Асимметричность мембраны. По химическому составу наружная поверхность мембран отличается от внутренней.
1) Мембраны ассиметричны по липидному составу. Существует асимметрия расположения фосфолипидов в мембране. Фосфатидилхолин и сфингомиелины локализованы в наружном слоя мембран, фосфатидилсерин и фосфатидилэтаноламин – во внутреннем. Холестерол находится преимущественно в наружном бислое.
2) Белки в мембранах расположены асимметрично (расположение периферических белков, различная степень погружения интегральных белков).
3) Наиболее асимметрично распределены в плазматической мембране гликолипиды и гликопротеины. Углеводные части гликолипидов и гликопротеинов выходят на наружную поверхность, иногда образуя сплошное покрытие на поверхности клетки – гликокаликс.
4. Динамичность мембран. Отдельные молекулы мембранных липидов и белков способны перемещаться в мембране, т.е. они сохраняют способность к диффузии. Молекулы белков и липидов с высокой скоростью двигаются в плоскости мембраны (латеральная диффузия). Они легко меняются местами со своими соседями в пределах одного монослоя примерно в 10 раз в секунду. Молекулы белков, так же как и липидов, способны к латеральной диффузии, однако скорость их диффузии в несколько раз ниже, чем молекул липидов. Перемещение мембранных белков в латеральной плоскости может быть ограничено вследствие притяжения между функционально связанными белками и образования кластеров, что в конечном счете приводит к их мозаичному распределению в липидном слое.
Кроме того, молекулы белков и липидов очень быстро вращаются вокруг своих продольных осей (вращательная диффузия).
Перескок липидных молекул из одного монослоя в другой (поперечная диффузия, flip-flop) осуществляется редко, а белки к такому перескоку вообще не способны. Причина исключительно медленного flip-flop заключается в его энергетической невыгодности, поскольку необходимо перенести полярную головку молекулы липида через гидрофобную область бислоя.
Дата добавления: 2015-06-12; просмотров: 4000;