ВВЕДЕНИЕ 3 страница
При обсуждении вопроса о локализации следов памяти следует иметь в виду, что в научении и воспроизведении прошлого события в большинстве случаев принимает участие не один какой-либо воспринимающий канал, например, зрение: чаще всего запоминаемый предмет мы видим, слышим и осязаем. Следует думать, что должны существовать связи между различными местами хранения следов отдельных элементов данного воспоминания. Связи эти, возможно, обеспечивают временную синхронизацию записанных в памяти следов при их воспроизведении. Это хорошо согласуется с результатами экспериментов, которые показывают, что некоторые следы памяти относительно хорошо сохраняются даже при обширных повреждениях мозга.
Интересные данные получены школой академика И. С. Бериташвили в экспериментах на собаках. У жи-
1 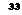 Л. М. Вейн, Б. И. Каменецкая
Л. М. Вейн, Б. И. Каменецкая
2 вотных производилось двустороннее удаление корковых областей слуховых и зрительных зон, связапных с анализом поступающей информации, и последующая выработка пищевых навыков. Опыты подтвердили роль корковых зон в процессах хранения следов памяти только па соответствующий раздражитель. Было показано также, что наиболее важное значение для сохранения в памяти воспринятых пищевых объектов имеют передпие отделы коры, точнее, лобные, или префронтальные, отделы коры. Двустороннее удаление их приводит к общим нарушениям образной памяти независимо от характера предъявленного раздражителя. Можно полагать, что височные доли мозга являются общей ассоциативной зоной для зрительного, слухового и вестибулярного восприятия, а также обонятельного и вкусового. Значение височных долей для функции памяти убедительно показал Пенфилд, раздражая их электрическим током.
Все эти данные определенно указывают иа существование специфических нейронных схем для записи прошлых событий.
Экспериментальные данные дают, кроме того, некоторые основания считать, что процессы обучения обеспечиваются, по-видимому, и стволовыми структурами мозга. Об этом свидетельствуют опыты Сперри на обезьянах с раздвоенным мозгом, которые должны были при появлении одного зрительного раздражителя тянуть па себя более шероховатый из двух рычагов, а при появлении другого — более гладкий рычаг. Обезьян приучали тянуть за рычаг не той рукой, которой управляем полушарие мозга, получающее зрительную информацию, а другой, контролируемой изолированным от всякой зрительной информации полушарием. Несмотря на полный разрыв прямых связей между полушариями, животные научались выполнять эту задачу, требующую координации различных раздражителей. Следовательно, осязательная и зрительная функции, осуществляемые разными полушариями, координировались каким-то подкорковым отделом, скорее всего на уровне ствола мозга. Поскольку структуры, несущие координирующую функцию, должны иметь тесные связи с каждой из специфических областей, участвующих в обучении, можно думать о ведущей роли в этом процессе ретикулярной формации верхних отделов ствола мозга.
Имеются также данные, которые позволяют считать, что следы менее сложных навыков локализуются, вероятнее всего, в стволе мозга, так как после удаления коры мозга (декортикации) у животных остается способность к выработке и сохранению следов. Известный латиноамериканский нейрофизиолог Э. Пеон установил, что кошка после удаления коры мозга может научиться поднимать лапу, чтобы избежать электрического удара после предупредительного звукового сигнала.
По-видимому, способность памяти фиксировать следы присуща вообще всем нейронам центральной нервной системы. Об этом свидетельствуют известные опыты американского исследователя Т. Чемберлена с обучением животных после удаления у них головного мозга (так называемым «спинальным» обучением). Исследователь показал, что удаление половины мозжечка вызывает возникновение четкой асимметрии позы задних конечностей. Важно, что существование вызванной таким путем асимметрии позы в течение определенного времени (от получаса до часа) сопровождалось стойким сохранением ее и после отделения спинного мозга от головного. Это значит, что в нейронах спинного мозга происходит фиксация каких-то функциональных сдвигов.
Спустя 5 лет аналогичные опыты произвели JI. П. Ла- таш и Л. И. Тихомирова, которые также выявили у оперированных животных четкую асимметрию спинальных рефлексов. Если асимметрия, как и в предыдущих опытах, существовала определенное время (свыше 20—30 минут), то последующая перерезка спинного мозга на границе грудного и поясничного отделов не приводила к ее исчезновению. Если же перерезку производили раньше, то она устраняла возникшую асимметрию ответов. Таким путем еще раз было показано, что нейроны спинного мозга способны фиксировать некоторые функциональные изменения и что для закрепления изменений на длительный срок необходим определенный интервал времени.
Мы изложили основные экспериментальные факты, касающиеся вопроса о том, где, в каких структурах мозга хранятся следы памяти. Далее встает вопрос о возможной локализации стойких изменений, лежащих в основе следа памяти, уже на клеточном уровне.
В настоящее время высказываются взгляды на неравнозначность для механизмов памяти нейронов коры мозга.
|
На основании изучения структурных особенностей нейронов С. А. Саркисов выдвинул положение об особой роли в следовых процессах звездчатых клеток коры больших полушарий, имеющих отношение к формированию образа.
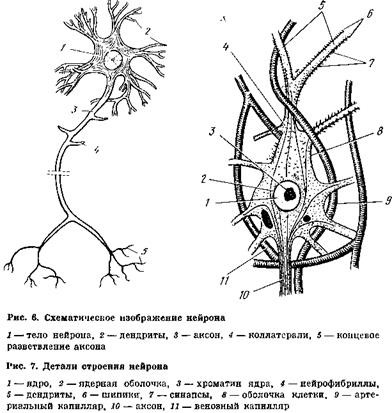
Хотя все исследователи согласны с тем, что формирование следа памяти связано с изменениями на нейронном уровне, однако пока еще не известно, ^акие структурные образования нейрона имеют к ним непосредственное отношение. Большинство ученых склонно думать, что основным элементом памяти является не весь нейрон в целом, а места его контакта с другими нейронами и их отростками — синапсы, расположенные па теле и на ко-
ротких ветвящихся отростках каждого нейрона, называемых дендритами (рис. 6, 7).
Это предположение кажется наиболее вероятным, так как среднее число синапсов на каждом нейроне может достигать тысячи. Следовательно, число элементов памяти в каждом нейроне огромно, что в большей степени может обеспечить требуемую емкость памяти.
Предположение о том, что основную роль в процессах памяти играют синапсы, подтверждается значительными изменениями в числе и расположении дендритов в процессе роста животных и человека, и особенно в первые годы жизни, когда приобретается огромный индивидуальный опыт. Сравнение нейронов и их отростков у человека в различном возрасте показывает, что значительно меняется количество дендритов в течение раннего периода развития. Относительный объем тел нейронов уменьшается, а объем дендритов увеличивается; поверхность дендритов возрастает, тогда как поверхность тел нейронов остается почти постоянной начиная с грудного возраста. Эти исследования, несомненно, указывают, что дендриты, разветвленность которых с годами сильно возрастает, имеют важное значение и, возможно, становятся базой для приобретения индивидуального опыта.
Близкие к этим данные получены при исследовании нервной системы в процессе эволюции животного мира — в филогенезе. Сравнительные исследования показывают, что структура нейрона и химические процессы в них меняются на протяжении эволюции очень незначительно. Основной признак, претерпевающий наиболее четкое развитие,— сложность и возрастающее количество связей между нейронами, а также увеличение их разнородности (гетерогенности). В настоящее время выявлено чрезвычайное разнообразие межнейронных связей и контактов между любыми частями нейронов. Особое значение имеют, по-видимому, шипики на дендритах. Эти образования представляют собой наиболее специализированные структуры, которые появляются только на определенном уровне эволюции мозга. Форма шипиков изменяется в связи с расширением и усложнением их функционального назначения. При определенных условиях в нервных центрах обнаруживается рост ответвлений и от длинного отростка нейрона — аксона. Возможно, что один из стимулирующих факторов роста — повышение функции в том или
другом нервном центре. По-видимому, так можно объяснить усиление синтеза белка и нуклеиновых кислот, в частности рибонуклеиновой кислоты — РНК — в тренируемых нейронах. (Более подробно на этих исследованиях мы остановимся в разделе о биохимических теориях памяти.) Если даже рост происходит со скоростью, не превышающей времени восстановления периферического нерва, то и тогда, как указывает Г. Д. Смирнов, за 10 минут могут образовываться волокна протяженностью 30 ммк. В масштабах организации нервных центров это величина значительная.
В объяснении характера происходящих изменений возможны три варианта: образование новых синапсов, включение пефункционирующих синапсов и изменение их функциональных свойств. Наиболее признаны теории, связанные с изменением функциональных свойств синапсов. Согласно теории проторения путей память объясняется повышением проводимости синаптических межнейронных контактов. Прохождение импульсов через синапсы облегчается тем, что импульсы уже ранее циркулировали по синаптическим путям. Это приводит к изменению физических, скорее всего электрических, характеристик синаптических пластинок. Изменение синапсов, их гипертрофия при функционировании доказаны экспериментально. Можно полагать, что гипертрофия синапсов сопровождается повышением их функциональной активности. Такого рода «синаптическое облегчение», по-видимому, лежит в основе и условных рефлексов. Некоторые нервные пути существуют от рождения.
Мозг состоит из нервных и глиальных клеток. В последние годы все более и более серьезное внимание обращается на роль клеток нейроглии в функциях мозга и, в частности, в процессах памяти. Этих клеток в 10 раз больше в мозгу, чем нейронов. Они занимают большую часть объема мозга и находятся в теснейшем соприкосновении с нейронами, заполняя промежутки между ними. Ранее им приписывалась лишь «подсобная» роль. В настоящее время накоплено много фактов, которые свидетельствуют об активном участии нейроглии в реакции мозга. По мнению американского нейрофизиолога Р. Галамбоса, глия «указывает нейронам порядок и последовательность их действия» и может влиять на их возбудимость. Учитывая способности глии размножаться и таким образом
увеличивать синтез белка, а также отвечать более длительными электрическими сдвигами, Галамбос приписывает нейронам функцию хранения информации.
Другие ученые приводят доводы в пользу того, что некоторые процессы переработки информации в мозгу обусловлены способностью глиальных клеток изменять проводимость электрического тока к отдельным участкам нейронов, в частности в области дендритов. Возможно, в этом и проявляется функция памяти.
С помощью микроэлектродной техники удалось, например, установить, что во время раздражения отдельных нейронов повышение синтеза РНК происходит в первую очередь в глиальных клетках и идет параллельно числу посылаемых импульсов. При использовании веществ, тормозящих синтез РНК, глиальный синтез уменьшается по сравнению с нейронным. На основании этих данных можно сделать вывод, что глиальные клетки более чувствительны к воздействиям, связанным с обучением. Опыты по содержанию животных в «обогащенной», т. е. богатой информацией, среде и в «обедненной» среде, почти лишенной какой-либо информации, выявили увеличение массы коркового вещества у животпых первой группы главным образом за счет элементов глии и подтвердили представления о повышепной их чувствительности.
Удалось также экспериментально показать, что импульсы возбуждения, приходящие в кору головного мозга, вызывают изменение активности не только нейронов, но также и клеток нейроглии. Аналогичные результаты получены американскими исследователями в опытах с внутриклеточным отведением биопотепциалов.
С функцией нейроглии связано образование миэлино- вых оболочек на волокнах центральной нервной системы, которые имеют огромное значение для эффективнрго проведения возбуждения через синапсы. На это указывают и данные филогенеза. У низших позвоночных, например у миноги, нейроглия слабо дифференцирована, и отсутствует миэлиновая оболочка на нервных волокнах. У этих животных нельзя выработать условных рефлексов, в то время как у костистых рыб условные рефлексы легко вырабатываются. Единственное отличие миноги от костистых рыб состоит в отсутствии дифференцированных клеток нейроглии и миэлиновой оболочки на нервных волокнах. Таким образом, появление в восходящем ряду позвоночпых способности к образованию условных рефлексов совпадает с дифференциацией нероглии.
Миэлинообразовательная деятельность клеток нейроглии, которая сохраняется и в зрелой нервной системе, стимулируется изменением заряда клетки нейроглии под влиянием нервного возбуждения. Об этом свидетельствуют двигательные реакции клеток, возникающие в течение секунд при добавлении в их среду ионов калия. Мгновенные двигательные реакции сопровождаются выпячиванием клеток нейроглии и ростом отростков, которые тут же закручиваются вокруг расположенных рядом с ними нервных волокон. Мгновенная реакция клеток нейроглии на приходящие нервные импульсы — важный объективный факт, подтверждающий предположение об участии нейроглии в осуществлении функции памяти. Изменение заряда в клетках нейроглии происходит скорее всего под влиянием ионов калия, выделяющихся из возбужденных нервных клеток. Есть основание полагать, что этот процесс происходит и под действием химических передатчиков нервного возбуждения — медиаторов, осуществляющих передачу нервного возбуждения. Последние выделяются отростками нервных клеток в области пресинапти- ческих окончаний, попадают в синаптическую щель и таким образом обеспечивают проведение нервных импульсов.
Согласно данным морфологии, пресинаптические волокна в коре головного мозга в основном «голые» и только 1/8 часть покрыта миэлином, который, благодаря более быстрому проведению возбуждения на этом участке, создает условия для прохождения большего количества электричества. В результате выделяется большее количество медиатора и, следовательно, обеспечивается лучшее функционирование синапса. Основываясь на изложенных фактах, тбилисский ученый А. И. Ройтбак выдвинул гипотезу, согласно которой немиэлинизированные синапсы не функционируют, а только отражают потенциальные возможности мозга. Включение их в нервную деятельность в процессе выработки условного рефлекса и обучения связано с активацией нейроглии. В результате происходят миэлинизация пресинаптических волокон и лучшее функционирование синапсов, принимающих участие в этом процессе.
Согласно этой иптересной гипотезе, возникшей па основе объективных данных, обучение сводится к процессу миэлинизации нервных волокон, который и обеспечивает фиксацию пережитого опыта. Такое представление, однако, несколько упрощает имеющиеся сложности в проблеме памяти и трудно согласуется с необычайной пластичностью нервной системы.
Мы рассмотрели современные нейрофизиологические представления, касающиеся процессов регистрации и хранения информации, возможной локализации следов памяти на макро- и микроскопическом уровне и подошли к самому трудному — к вопросу о механизмах воспроизведения информации.
Механизм воспроизведения, осуществляющий организацию поиска следа и извлечение хранимой информации, пока еще мало изучен. Приведенные выше данные о чрезвычайной прочности долговременных следов памяти позволяют предполагать, что нарушения памяти чаще всего обусловлены невозможностью их извлечения. Об этом же свидетельствует наблюдаемое нередко в повседневной жизни временное забывание отдельных событий. По-видимому, успех процесса воспроизведения в значительной степени обусловлен нормальной организацией запечатле- пия следов. Вследствие же его нарушения иногда невозможно найти «адрес хранения» следа и его извлечение.
Клинические наблюдения и данные эксперимента заставляют считать, что в осуществлении процесса воспроизведения принимают участие глубинные лимбико-ретикулярные структуры мозга, т. е. те же структуры мозга, которые имеют непосредственное отношение и к регистрации следов памяти. Так, у больных с двусторонним поражением структур гиппокампова круга помимо нарушения кратковременной памяти, как правило, наблюдается выпадение памяти на прошедшие события — ретроградная амнезия, которая представляет собой в основном нарушение процессов воспроизведения. Этот вид амнезии охватывает события, которые были нормально восприняты, длительно сохранялись и даже повторно воспроизводились. Следовательно, в данном случае речь может идти скорее всего о нарушении воспроизведения событий, поскольку их сформированные следы сохраняются в мозгу, по-видимому, всю жизнь. Прямым доказательством высказанного положения является обратимость ретроградной амнезии с полным восстановлением забытых событий.
Анализ клинических фактов свидетельствует об определенной роли структур гиппокампова круга в процессах воспроизведеыпя.
Эксперименты американского нейрофизиолога Р. Бикфорда и его сотрудников с электрической стимуляцией глубинных отделов мозга, расположенных в височной доле, выявили у больных аналогичные нарушения памяти па прошедшие события, которые также носили обратимый характер и, следовательно, были связаны с расстройством воспроизведения. При этом установлено, что, чем длительнее раздражение височных структур, тем продолжительнее период амнезии. Так, воздействие тока в течение одной секунды вызывало выпадение памяти на несколько минут, в то время как 10-секундная стимуляция нарушала память на несколько недель. Эти данные показали прямую зависимость между длительностью выключения височных структур мозга и продолжительностью нарушенного воспроизведешш. Они явились убедительным подтверждением клинических наблюдений о роли глубинных структур височных долей в организации процесса воспроизведения. Тем не менее объяснение полученных данных представляет значительные трудности, поскольку воспроизведение событий нарушается в течение ограниченного периода времени. В связи с этим нельзя говорить о расстройстве каких-то общих механизмов воспроизведения.
При обсуждении вопросов, касающихся процесса воспроизведения, особый интерес представляют упомянутые выше исследования У. Пенфилда, который при раздражении электрическим током корковых областей височных долей мозга вызывал яркие воспоминания о прошлом. Исследования Пенфилда с несомненностью указывали на связь этого явления с активацией следов памяти и вызвали большой интерес к функциональной значимости корковых областей височной коры. Более детальный анализ наблюдения показал, что воспоминания прошлого возникали при раздражении нижнего края одной из височных долей только у больных, страдающих эпилептическими припадками. Опыты Пеифильда дают основание говорить о возможности возникновения этого явления лишь при снижении нормального порога возбудимости, и в частности снижения порога активации следов памяти. Пенфилд отмечал, что при височной эпилепсии яркие воспоминания 42 прошлого возникали во время припадка, обусловленного поражением тех же височных структур, без их искусственной стимуляции.
Вызванные таким путем воспоминания, как правило, воспроизводили реальные события прошлого. Но обычно это были воспоминания, на которых больной не фиксировал свое внимание: воспроизводимые эпизоды чаще всего не содержали в себе ничего значительного. Возможность воспроизводить через длительное время как будто бы прошедшие мимо и не запомнившиеся эпизоды прошлого со всеми их незначительными подробностями позволяет думать, что в мозгу записываются и хранятся все события, которые проходят перед нами. Во всяком случае, хранится значительно больше того, что мы способны произвольно вспомнить.
Воспроизведение событий прошлого, вызванных электрической стимуляцией, характеризуется яркостью; этим они отличаются от образов обычной памяти. По Пенфил- ду, «больные никогда не рассматривали возникавшие переживания как воспоминание. Скорее, это значило для них слышать или вновь видеть, т. е. снова переживать мгновения прошлого». Тем не менее больной не теряет при этом связи с настоящим. Важно отметить, что раздражение других корковых зон, помимо височной, не приводило к подобному эффекту. Тот факт, что определенные корковые отделы височной доли — единственная область коры, электрическое раздражение которой вызывает образы памяти, позволил думать об особом ее функциональном назначении в процессе воспроизведения. По- видимому, эта область мозга — единственный корковый путь к следам прошлых событий, и, возможно, к записи следов памяти. Из полученных данных не следует, однако, заключение о том, что запись событий должна происходить именно в височной части коры, так как удаление одной височной доли не вызывает расстройств памяти.
Для процесса воспроизведения важное значение имеет функциональное состояние нервной системы. В этом отношении представляет большой интерес явление так называемого диссоциированного обучения. Оно заключается в том, что у животных при различном состоянии центральной нервной системы могут быть выработаны независимые условные рефлексы. Успешное припоминание навыка становится возможным только в том состоянии мозга, в котором производилась их выработка. У животного возникают как бы разобщенные состояния, при которых воспроизводятся различные условные рефлексы: проявление условного рефлекса в одном состоянии не влияет на воспроизведение его в другом.
Это явление, изучаемое А. Н. Черкашиным и его сотрудниками в Институте биофизики АН СССР, можно продемонстрировать в следующем эксперименте. Крыс в нормальном состоянии обучали передвигаться в лабиринте в правую камеру, а в левую — после введения некоторых химических веществ, например, аминазина, изменяющего функциональное состояние мозга. Химическое вещество переводило животное в новое функциональное состояние и устраняло ранее выработанные навыки.
С точки зрения диссоциированного обучения можно, по-видимому, понять нарушение репродукции хорошо заученных фактов в состоянии сильного эмоционального возбуждения, обусловленного влиянием на центральную нервную систему гормональных факторов, изменяющих функциональное состояние мозга. Факт диссоциированного обучения находится в некотором противоречии с многочисленными данными о сохранении условных рефлексов, выработанных под влиянием различных фармакологических веществ, даже после окончания их действия.
Необходимо дальнейшее изучение этого интересного явления, которое является хорошей моделью для анализа механизмов записи и считывания информации, а также влияния на них различных химических веществ.
Уровень бодрствования несомненно оказывает влияние на отдельные фазы памяти, в том числе и на процесс воспроизведения. Известно, что для восприятия необходим какой-то минимальный уровень бодрствования: в период сна и в бессознательном состоянии восприятие невозможно. Нормальное протекание переработки уже воспринятой информации не зависит от состояния сна и бодрствования. Напротив, оптимальные условия для воспроизведения наблюдаются при низком уровне бодрствования или при его отсутствии: во сне, гипнозе, во время барбитурового наркоза.
Не менее важным фактором, влияющим на память, и в частности на процесс воспроизведения, как уже указывалось, является состояние эмоциональной сферы. Практика показывает несомненную роль эмоций в процессах обучения и запоминания.
Хорошо известно, что вся поступающая в течение жизни информация не может быть усвоена и извлечена из памяти, а оперируем мы только тем, что имеет определенную значимость. Такая сортировка и отбор информации по значимости обеспечиваются включением аппарата эмоций. Клинические исследования подтверждают существующую в норме тесную функциональную связь памяти и эмоций и дают яркие примеры нарушений памяти, возникающих на базе первичных эмоциональных расстройств.
Следует подчеркнуть, что состояние эмоциональной сферы имеет значение и для процесса воспроизведения. Хорошо известно, что выраженное эмоциональное напряжение может приводить к нарушению воспроизведения, например к забыванию во время экзамена или ответственного выступления усвоенных знаний, которые вспоминаются немедленно, как только человек приходит в спокойное состояние. В то же время известны случаи, когда волнение позволяет в ответственный момент вспомнить как будто бы забытые факты.
Приведенные данные, несомненно, свидетельствуют о том, что для нормального осуществления процесса воспроизведения необходимо участие целого ряда структур и функциональных систем, в том числе необходимо участие механизмов, лежащих в основе бодрствования, внимания, эмоций и оказывающих влияние на функциональное состояние мозга. Все это указывает на чрезвычайную сложность процессов воспроизведения и функции памяти в целом, в которых принимают участие перечисленные системы мозга. Последние вплетаются в организацию этой комплексной функции и помогают ее осуществлению.
При обсуждении физиологических теорий памяти несомненный интерес представляют исследования академика М. Н. Ливанова и его сотрудников, отражающие современные представления нейрофизиологии об организации мозговых функций.
М. Н. Ливанов на основании многолетнего изучения этой проблемы приходит к выводу, что в процессе обучения возникают новые системы реагирования нейронов, в основе которых лежит изменение условий, обеспечивающих проведение возбуждения в коре головного мозга. Удалось установить, что к таким условиям относятся син- хроиность процессов в различных участках коры, определенная частота колебаний биоэлектрических потенциалов и совпадение их по фазам. Иными словами, если возбудимость повышена в одном участке коры, то для проведения возбуждения она должна быть повышена и в другом соответствующем ей участке. По-видимому, эти условия мгновенно возникают под влиянием условного сигнала и также мгновенно распадаются, а кора остается свободной для всех форм деятельности. Следовательно, пет надобности связывать обучение с возникновением фиксированных рефлекторных дуг. Результаты исследований М. Н. Ливанова дают основание считать, что процессы памяти, в частности условный рефлекс, который представляет простую форму памяти, формируются на уровне функциональной системы, а кодирование и удержание информации идут, по-видимому, на уровне синаптических аппаратов. Приведенные соображения подчеркивают роль функциональной системы в процессе запоминания и обучения и не позволяют свести его только к изменениям отдельных нейронов даже при низких формах организации нервной системы.
Биохимические основы памяти
Если в физиологических теориях ученые пытаются подойти к механизмам памяти на уровне функциональных систем и дают анализ различных фаз памяти, то сторонники биохимических теорий, стремясь выявить механизмы, связанные с формированием следа долговременной памяти, адресуются главным образом к процессам хранения следа на нейронном уровне. Согласно биохимическим теориям основное значение в хранении мозгом информации принадлежит изменениям химического состава нуклеиновых кислот и белков соответствующих нейронов. Такие представления возникли в связи с достижениями в расшифровке механизмов наследственной памяти.
Сразу же сделаем оговорку: генетическая информация — видовая память — передается от клетки к клетке, от организма к организму, т. е. она наследуется. Индивидуальная память не наследуется. Наш опыт, все то, чему мы научились, невозможно передать нашим детям, Они должны всему учиться сызнова.
Биохимикам и генетикам удалось доказать, что передача информации по наследству осуществляется с помощью дезоксирибонуклеиновой — ДНК — и рибонуклеиновой — РНК — кислот. Это натолкнуло биохимиков на мысль, что, может быть, и индивидуальная память осуществляется за счет таких же механизмов. В пользу подобного предположения говорит и огромная информационная емкость нуклеиновых кислот, которая может обеспечить храпение большого количества информации. В отличие от генетической памяти, главная роль в процессах индивидуальной памяти приписывается молекулам РНК.
Большая работа в этом направлении проведена известным шведским ученым Г. Хиденом, который разработал методику изъятия из мозга животных отдельных нервных клеток и определения в них количественного и качественного состава РНК. Обучая крыс определенным формам поведения, Хиден наблюдал заметное увеличение количества РНК в нервных клетках, участвующих в выработке вновь приобретенных навыков. Так, если животное передвигалось по натянутой проволоке, то в нервных клетках вестибулярного аппарата, осуществляющего равновесие, возрастало количество РНК. При обучении крыс доставать передней лапой пищу количество РНК увеличивалось в клетках коры головного мозга, ответственных за регуляцию движений именно этой конечности.
В 1959 г. Хиден сообщил, что в клетках мозга обученных крыс увеличивалось содержание РНК на 12% по сравнению с клетками мозга крыс, живущих в нормальных условиях. Когда крысу-левшу научили пользоваться правой лапой, чтобы доставать корм из трубки, клетки в коре мозга вырабатывали не только белки, но и особый вид РНК. То же самое наблюдалось и у золотых рыбок, которых заставляли освоить новый стиль плавания, прикрепляя им к нижней части головы плавучий пенопласт. У рыбок, выполнявших не менее тяжелую работу, когда их заставляли плыть против течения, выработки дополнительной РНК не отмечалось.
Согласно гипотезе Хидена, нервное возбуждение изменяет состав РНК, что в свою очередь ведет к образованию белка, соответствующего этому возбуждению. Таким образом, под влиянием нервного возбуждения может возникнуть новый белок, являющийся материальным следом, отпечатком, в котором отложен опыт. Когда же снова возникает сходное возбуждение, оно как бы активирует этот белок. Так был получен первый намек на участие РНК в процессах обучения.
Дата добавления: 2015-02-16; просмотров: 723;