ГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ РАЗНЫХ ФОРМ ДВИГАТЕЛЬНОЙ
активности дрозофилы. суточный ритм. Для анализа общебиологических закономерностей контроля жизнедеятельности не только насекомых, но и других групп животных представляют интерес генетические изменения суточного ритма. Исследуются, например, мутации дрозофилы, изменяющие длительность циркадного (суточного) ритма уровня двигательной активности мух и ритма массового вылупления взрослой формы из куколки. При мутации в локусе arythmic суточные колебания активности отсутствуют; мутации в локусе period, либо укорачивают цикл активности (short period) до примерно 19 ч, либо удлиняют его (long period) до примерно 28 ч. При мутации per 0 суточный ритм практически отсутствует.
У разных видов дрозофилы описано около 10 мутаций, влияющих на суточный ритм. Они обнаруживаются как в их воздействии на суточные колебания локомоторной активности, так и в модуляции периода массового вылета взрослых особей из куколок. Ритмичность физиологических процессов, влияющих на локомоцию и приуроченность вылупления к определенной фазе циркадного ритма, обусловлена согласованием световосприятия с работой эндогенного водителя ритма (пейсмейкера).
Это было показано при исследовании аутосомной рецессивной мутации lime, когда ритмичность вылупления при определенных условиях поддерживается, однако смена света и темноты не может запускать ритм.
Геотаксис. Первые систематические и подробные эксперименты по генетическому контролю геотаксиса у Dr. melanogaster провел в 60-е годы Дж. Хирш (см.: Эрман и Парсонс, 1984). Геотаксис, т.е. перемещение мух. ориентированное относительно силы тяжести, оценивали с помощью специально сконструированных лабиринтов (рис. 8.10).

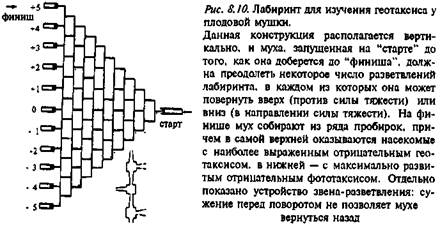 Использовали вертикально расположенный лабиринт, представляющий собой систему соединенных друг с другом модулей, в каждом из которых муха может выбрать между поворотом в одну сторону, который ведет ее вверх, и поворотом в другую, при котором она спускается вниз. Обычно использовали 10- или 15-модульный лабиринт. Группу мух помещали в единственную пробирку, расположенную на стартовой стороне лабиринта, и давали им возможность перемещаться в направлении другой стороны лабиринта. Привлекательными стимулами были либо запах пищи, либо свет (рис. 8.10). Поскольку на финишной стороне лабиринта находится не одна, а 10 или 15 пробирок, расположенных на разной высоте по отношению к стартовой, то происходит сортировка мух по уровню проявления их геотаксиса. Такая методика позволяет провести отбор на положительный и отрицательный геотаксис у дрозофилы (рис. 8. 11), причем селекция происходит очень быстро. В дальнейшем было проведено аналитическое исследование роли "вклада" генов каждой из хромосом дрозофилы (за исключением 4-й) в обеспечение отрицательного и положительного геотаксиса. Это оказалось возможным благодаря использованию специальных линий, которые несли доминантные маркеры, а также инверсии, препятствовавшие кроссинговеру определенных хромосом. Была получена количественная характеристика линий дрозофилы, селектированных на разные уровни геотаксиса, и определена степень участия разных хромосом в детерминации этого признака (подробнее см.: Эрман, Парсонс, 1984).
Использовали вертикально расположенный лабиринт, представляющий собой систему соединенных друг с другом модулей, в каждом из которых муха может выбрать между поворотом в одну сторону, который ведет ее вверх, и поворотом в другую, при котором она спускается вниз. Обычно использовали 10- или 15-модульный лабиринт. Группу мух помещали в единственную пробирку, расположенную на стартовой стороне лабиринта, и давали им возможность перемещаться в направлении другой стороны лабиринта. Привлекательными стимулами были либо запах пищи, либо свет (рис. 8.10). Поскольку на финишной стороне лабиринта находится не одна, а 10 или 15 пробирок, расположенных на разной высоте по отношению к стартовой, то происходит сортировка мух по уровню проявления их геотаксиса. Такая методика позволяет провести отбор на положительный и отрицательный геотаксис у дрозофилы (рис. 8. 11), причем селекция происходит очень быстро. В дальнейшем было проведено аналитическое исследование роли "вклада" генов каждой из хромосом дрозофилы (за исключением 4-й) в обеспечение отрицательного и положительного геотаксиса. Это оказалось возможным благодаря использованию специальных линий, которые несли доминантные маркеры, а также инверсии, препятствовавшие кроссинговеру определенных хромосом. Была получена количественная характеристика линий дрозофилы, селектированных на разные уровни геотаксиса, и определена степень участия разных хромосом в детерминации этого признака (подробнее см.: Эрман, Парсонс, 1984).
Поведение мух, селектированных на отрицательный геотаксис в течение 181 поколения, имело целый ряд особенностей, привнесенных процедурой тестирования в лабиринте. Оказалось, что параллельно с отбором на крайние значения геотаксиса эти мухи подвергались неосознанному отбору: на устойчивость к высыханию и способность переносить длительные периоды голодания, поскольку перемещение некоторых мух занимало до нескольких часов и многие из них погибали; на сниженный уровень двигательной активности, поскольку более активные насекомые имели тенденцию к гибели; на усиление стереотипии в поведенческих реакциях, поскольку мухи, обладающие крайними значениями геотаксиса, имели тенденцию делать повороты в одну и ту же сторону, в том числе и в лабиринте, который был расположен горизонтально; на сниженный уровень "клаустрофобии", поскольку мухам приходилось преодолевать суженные участки пробирок при переходе из одного блока лабиринта в другой и некоторые насекомые не справлялись с этим.
Результаты экспериментов показывали сложность поведения дрозофилы и возможность формирования иных, чем предполагалось изначально, ответов на отбор. Подобное явление относится к так называемым коррелированным ответам, достаточно подробно изученным также в экспериментах по генетике поведения грызунов (см.: 8.6.3.1).
Фототаксис.Следующий этап в накоплении информации о генетических основах поведения дрозофилы связан с именем американского ученого С. Бензера (1975), в работах которого был использован целый комплекс экспериментальных подходов.
В частности, был активизирован поиск неврологических мутантов, и с этой целью были разработаны специальные приемы массового тестирования мух.
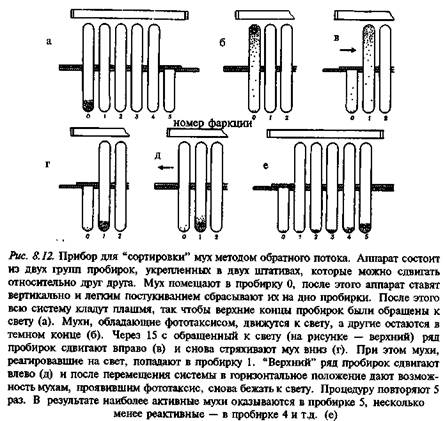
Отбор потенциальных носителей аномалий проводили с помощью метода обратного потока (рис. 8.12). По выражению Бензера, этот способ позволял сортировать мух, как молекулы. Сущность его заключалась в следующем. Разделяли мух, которые двигались внутри трубки в направлении источника света, и тех, которые оставались на месте. Процедура повторялась несколько раз. В результате была получена серия групп насекомых, у которых выраженность исследуемого признака варьировала от минимального до максимального значения.
На следующем этапе из числа мух с положительным фототаксисом следовало отделить более активных особей от тех, которые энергичнее шли именно на свет. Мух с отрицательным фототаксисом также разделяли на малоактивных и на тех, которые избегали света. Сэтой целью мухам предлагал ось двигаться в трубке от источника света. В результате можно было получить 5 групп насекомых, 4 из которых попарно различались либо по знаку фототаксиса, либо по уровню активности, у 5-й группы фототаксис отсутствовал (они не реагировали на свет) и были малоактивными (рис. 8.13). Полученные группы насекомых, достоверно различавшиеся по поведению, можно было исследовать нейрофизиологически. Запись электрических потенциалов глаза показала, что у части особей электроретинограмма отсутствовала. В ряде случаев у молодых мух электрическая реакция глаза на свет была нормальной, но затем происходила дегенерация фоторецепторов.
 Особенности формирования аномального фенотипа у мутантов с нарушенным зрением были исследованы в экспериментах с гинан-дроморфными мозаиками (см.: 8.5.3.2). Первой в этом ряду была классическая работа А. Хотта и С. Бензера, кратко изложенная выше.
Особенности формирования аномального фенотипа у мутантов с нарушенным зрением были исследованы в экспериментах с гинан-дроморфными мозаиками (см.: 8.5.3.2). Первой в этом ряду была классическая работа А. Хотта и С. Бензера, кратко изложенная выше.
Проявление признака – аномальные зрительные реакции – исследовали у особей, часть клеток которых не имела Х-хромосомы. В таких клетках (они развились по "самцовому" типу) обнаруживался эффект упомянутых выше рецессивных генов, влиявших на фенотип.
Судорожные состояния.Еще одна группа аномалий с мутациями в Х-хромосоме была обнаружена случайно при наркотизации мух в ходе очередной серии опытов. Оказалось, что у этих особей при действии эфирного наркоза развиваются быстрые подрагивания конечностей или вспышки некоординированных движений, которые распространяются среди группы мух наподобие цепной реакции. Наиболее полно описаны мутации shaker (sh) и hy-perkinetic (hk), в частности выявлены аллели гена hk, при которых мухи резко подпрыгивают и падают на спину при внезапном появлении в поле зрения движущегося предмета.
В знаменитых экспериментах американских ученых К. Икеды и В. Каплана в начале 70-х годов с помощью метода гинандроморф-ных мозаиков был прослежен путь проявления таких аномалий. Анализ физиологических процессов у особей, ткани которых представляли собой "мозаику" из участков нормального и мутантного генотипов, показал, что подергивание лапок при эфирном наркозе наблюдается не у всех мух. Мотонейроны, отвечающие за движения конечностей, располагаются в торакальном ганглии. Если ткань этого ганглия оказывается полностью или частично мутантной, то движения всех или части конечностей аномальны. Если же ткань торакального ганглия не несла мутантного аллеля, то аномальных подергиваний конечностей не было, даже если выше расположенные участки ЦНС (надглоточный ганглий) были мугантными. Таким образом, для проявления эффекта мутации принципиально важным оказывается экспрессия мутантного аллеля в нервных клетках, непосредственно проецирующихся в мышцы (рис. 8.14).
8.4.3.5. МУТАЦИИ, ВЛИЯЮЩИЕ НА ПОЛОВОЕ ПОВЕДЕНИЕ.У большин-
ства видов животных собственно копуляции предшествует ритуал ухаживания, в процессе которого партнеры по очереди осуществляют видоспецифические движения (ФКД). Половое поведение дрозофилы намного примитивнее, чем у птиц и млекопитающих, коммуникационные процессы которых трудны для исследования именно из-за их сложности. У дрозофилы простота и четкость взаимодействий партнеров при спаривании видоизменяются в результате некоторых мутаций, что позволяет уточнить сложную мозаику их чередования.
Уже упоминалось о влиянии мутации yellow на фазу вибрации при ухаживании самца: она оказывается укороченной, и уровень стимуляции самки, необходимый для продолжения ритуала ухаживания,  достигается только потому, что возбудимость самок этой линии выше, чем у самок дикого типа. Нарушение нормального ритма вибрации происходит и при мутации cacophony.
достигается только потому, что возбудимость самок этой линии выше, чем у самок дикого типа. Нарушение нормального ритма вибрации происходит и при мутации cacophony.
Частота биений крыла, длительность отдельных "пульсов" и интервалов между ними – это признаки, по которым различаются близкие виды, обеспечивая межвидовую изоляцию. Так, например, виды-двойники – Dr.simulans и Drjacuba – различаются "рисунком" видовой "песни"-вибрации. Поскольку движения крыла при вибрации – это пример примитивного инстинктивного движения, по своим свойствам относящегося к ФКД, генетические различия подобного рода подтверждают предположение о существовании резерва генетической изменчивости, обеспечивающего варьирование частоты выполнения ФКД (см.: 8.2.5).
Генетический контроль ухаживания очевидным (а может быть, и тривиальным) образом связан с процессом половой дифференци-ровки, поскольку мутации, затрагивающие последнюю, влияют и на поведение ухаживания. Выше упоминалось о том, что после спаривания в половых путях самки образуется специальное сигнальное вещество – антиафродитный феромон г<мс-вакцинилацетат. Процесс его формирования сложный. Оказалось, что образование сигнальных веществ этой группы связано с генотипом самца, спаривание с которым было успешно завершено. В сперме самца присутствует специфический фермент из группы эстераз, конкретная изоферментная форма которого детерминируется определенным ал-лелем данного локуса.
Именно его каталитическая активность в половых путях самки продуцирует (или не продуцирует) г<ис-вакцинилацетат.
Роль поведения самки в осуществлении самцом ритуала ухаживания была проанализирована еще в 70-е годы с использованием метода гинандроморфных мозаиков (см.: 8.5.3.2). Половое поведение мозаичных по "составу" своего тела мух, у которых доли "самцо-вых" и "самочьих" тканей различались, в разных условиях варьировало. Однако для процедуры простого преобладания "самочьих" черт строения было недостаточно. Критическими в этом отношении оказались определенные участки тела, а именно спинные хитиновые щитки – тегриты, щитки у основания ног – стерниты, а также область грудного интегумента (покрова). Когда эти участки имели генотип самки, то, даже если по другим морфологическим признакам муха была похожа на самца, она вызывала у самцов четкую реакцию ухаживания. Было показано, что ухаживание, в частности фаза вибрации, запускается обонятельными ключевыми раздражителями (феромонами), источником которых и являются указанные участки тела.
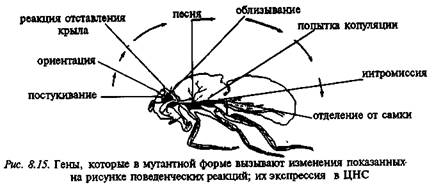
Исследование полового поведения гинандроморфных мозаиков показало, что эффекты разных мутаций определяются изменениями в разных участках ЦНС мухи. Отставление крыла и ориентация самца относительно самки, нарушающиеся при некоторых мутациях, связаны с работой надглоточного ганглия. Она повреждается при мутациях fruitless и celibate, и это ведет к нарушению копуляции. Изменение видоспецифической "песни"-вибрации при мутации cacophony связано с дефектом в торакальном ганглии мухи. О сложности механизма ритуала ухаживания говорит и тот факт, что он изменяется при мутациях в локусе per, когда нарушается и цир-кадная ритмика ряда процессов (см. выше). Возможно, что физиологические механизмы, обеспечивающие ритмические осцилляции, и механизмы, координирующие последовательности действий при ухаживании, либо управляются из одного отдела ЦНС, либо связаны с функцией одной медиаторной системы (рис. 8.15).
8.4.3.6. мутации обучения и памяти у дрозофилы.
Для массового тестирования поведения мух широко используется метод "обратного потока" (см. рис. 8.12). Несложное техническое устройство, которое позволяет "фракционировать" мух с разной выраженностью соответствующего признака, используется практически во всех исследованиях, связанных с обучением дрозофилы (рис. 8.16).

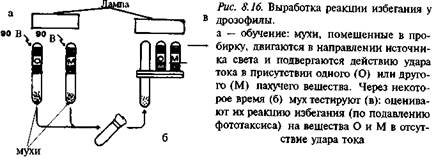 Группу из 30–50 мух помещают в специальную трубку, внутри которой они могут перемещаться в направлении, например, источника света. В тестовую трубку подают пары вещества, обладающего сильным, но индифферентным для них запахом. Продувание паров пахучего вещества сопровождается подачей электрического тока на специальные металлические пластины-электроды, протянутые вдоль всей трубки. Включение тока вызывает у мух оборонительные реакции. По прошествии определенного интервала времени их можно тестировать повторно, предлагая на этот раз только запах, но не включая электрический ток. При этом регистрируют число мух, которые все-таки переместились в направлении света при предъявлении условного (запахового) сигнала, несмотря на то что ранее она сопровождалась неприятным стимулом. Это число мух сопоставляется с данными по той же группе насекомых, когда они просто перемещались в градиенте нового для них, но также нейтрального запаха.
Группу из 30–50 мух помещают в специальную трубку, внутри которой они могут перемещаться в направлении, например, источника света. В тестовую трубку подают пары вещества, обладающего сильным, но индифферентным для них запахом. Продувание паров пахучего вещества сопровождается подачей электрического тока на специальные металлические пластины-электроды, протянутые вдоль всей трубки. Включение тока вызывает у мух оборонительные реакции. По прошествии определенного интервала времени их можно тестировать повторно, предлагая на этот раз только запах, но не включая электрический ток. При этом регистрируют число мух, которые все-таки переместились в направлении света при предъявлении условного (запахового) сигнала, несмотря на то что ранее она сопровождалась неприятным стимулом. Это число мух сопоставляется с данными по той же группе насекомых, когда они просто перемещались в градиенте нового для них, но также нейтрального запаха.
Достоверность различий в реакциях на запах с "наказанием" и на истинно нейтральный запах (получавшие удар тока мухи двигались к свету медленнее) свидетельствовала об имевшем место научении насекомых.
При описании эффектов наиболее известных мутаций, вызывающих аномалии обучения и памяти, таких как dunce (dnc), turnip (tur), cabbage (cab), rutabaga (rut), zuccini (zuc), локализованных в Х-хромо-соме, было продемонстрировано, что процесс восприятия запаха у таких мух не изменен, т.е. сенсорная функция не нарушена, следовательно, дефект связан с нарушением других функций ЦНС.
При мутации dnc резко сокращается время существования следа краткосрочной памяти. Через 10–15 секунд после сочетания стимулов эти мухи уже "не помнят", что получили наказание. У мутантов rut изменена не только краткосрочная память, но страдает и сам процесс обучения. Мутации tur и amnesiac (amn) также характеризуются сильно нарушенной памятью, хотя и в меньшей степени, чем dnc.
Полученные в экспериментах мутации, влияющие на процесс обучения реакции избегания удара тока и на хранение следов памяти, были в дальнейшем проанализированы с точки зрения изменений естественного поведения мух. Напомним, что в нормальных условиях самец дрозофилы делает попытку ухаживать за любыми самками, в том числе и за невосприимчивыми к ухаживаниям после оплодотворения. Последние на попытку уха живания отвечают, как уже отмечалось, специфической реакцией выделения антиафродитного феромона, что время после сеанса бучения, мин вызывает у самца длительное (1–2 ч) подавление готовности к спариванию даже с рецептивными самками. Оказалось, что самцы с мутацией dnc готовы ухаживать за любыми самками (и даже за самцами) сразу же после контакта с оплодотворенной самкой. Мухи с мутацией атп, при которой дефект памяти незначителен, "помнят" об опыте отрицательного взаимодействия несколько дольше. Мутация атп нарушает также готовность самок к спариванию (рис. 8.17).
Исследование нейрохимических коррелятов обучения и памяти у дрозофилы послужило мощным толчком к большой серии работ, посвященных роли генетического аппарата клетки в формировании следа памяти. На первых этапах такого исследования было выявлено, что данная группа мутационных изменений поведения характеризуется нарушениями в работе системы вторичных посредников, а именно в ферментной системе синтеза и деградации циклического аденозинмонофосфата, а также ферментных систем, связанных с последующим фосфорилированием белков.
Было показано, что ген dnc – это структурный ген фосфодиэсте-разы II дрозофилы. Найдена гомология между нуклеотидной последовательностью этого гена и последовательностью, кодирующей белок-предшественник гормона откладки яиц аплизии (см.: 8.3.4). Этот факт интересно сопоставить с описанной для мутантов по dnc стерильностью самок, когда образование яиц сохраняется, но самка не может их отложить.
Далее было продемонстрировано, что при мутации в локусе rut нарушено свойство аденилатциклазы активировать кальций калмо-дулина. При мутации tur ослабляется активность протеинкиназы С, а также гуанилатциклазы с одновременным снижением числа рецепторов моноаминов. При мутации атп повышается уровень цАМФ.
Участие системы вторичных посредников в формировании следа памяти обнаружено у аплизии, дрозофилы и млекопитающих (у мышей).
Общая схема влияния системы вторичных посредников на формирование памяти достаточно сложна, но очень упрощенно ее можно представить следующим образом. Внутриклеточный уровень цАМФ формируется вследствие активации аденилатциклазы под действием нейромедиатора. Циклический монофосфат, связываясь с протеин-киназой А, вызывает высвобождение каталитических субъединиц этого фермента, которые уже в ядре фосфорилируют так называемый белок CREB (cAMF response element binding protein) в положении Ser-133. CREB относится к категории регуляторных белков – транскрипционных факторов. В свою очередь он активирует ряд генов. Система активации CREB является звеном в цепи событий, участвующих в сохранении следа памяти в нервной системе апли-зии, долговременной памяти дрозофилы (Yin et al., 1994) и мыши (Bourchaladze et al., 1994; см. также: 8.6.3.3).
8.5.
Дата добавления: 2015-01-10; просмотров: 1513;