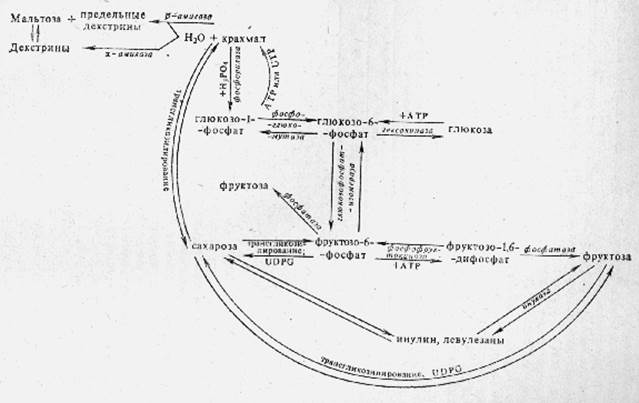
Образование и взаимные превращения углеводов
Первичным продуктом фотосинтеза является фосфоглицериновая кислота. При дальнейших превращениях она дает различные моносахариды - глюкозу, фруктозу, маннозу и галактозу. Эти моносахариды образуются без участия света, исключительно в результате «темновых» ферментативных реакций. Образование гексоз из фосфоглицериновой кислоты или фосфоглицеринового альдегида происходит благодаря действию фермента альдолазы. Этот фермент катализирует реакцию взаимодействия фосфоглицеринового альдегида и фосфодиоксиацетона с образованием фруктозодифосфата: Фосфодиоксиацетон + 3-фосфоглицериновый альдегид ↔ фруктозо-1,6-дифосфат. Альдолаза широко распространена в растительном мире, в наибольшем количестве содержится в активно растущих частях растений.
Наряду с моносахаридами в листьях чрезвычайно быстро образуются также сахароза и крахмал. Однако образование сахарозы и крахмала - вторичный процесс ферментативных превращений ранее образовавшихся моносахаридов.
Взаимопревращения моносахаридов происходят в результате действия соответствующих ферментов, катализирующих реакции фосфорилирования и образования фосфорных эфиров сахаров. Разнообразные гексозофосфорные эфиры найдены в целом ряде растений. Так, например, под действием фермента гексокиназы глюкоза превращается в глюкозо-6 фосфат. Под действием фермента глюкозофосфатизомеразы происходит обратимое превращение глюкозо- 6-фосфата во фруктозо-6-фосфат и маннозо-6-фосфат. В результате действия фосфоглюкомутазы глюкозо-6-фосфат может обратимо превращаться в глюкозо-1-фосфат. Фосфофруктокиназа катализирует превращение фруктозо-6-фосфата во фруктозо-1,6-дифосфат.
В растениях найдены также изомеразы, катализирующие взаимопревращения уроновых кислот, а также ксилозы и арабинозы. Образование свободных моносахаридов из их фосфорных эфиров происходит под действием фосфатаз, широко распространенных в растениях.
Чрезвычайно широко распространена в растениях сахароза - углевод, синтезирующийся только в растительном организме и играющий очень большую роль в обмене веществ у растений. В некоторых растениях сахароза может накапливаться в чрезвычайно больших количествах, например в сахарной свекле и сахарном тростнике. Синтез сахарозы в растениях происходит путем реакции трансгликозилирования с участием фермента сахарозосинтазы и уридинфосфатов фруктозы или фруктозо-6-фосфата.
Синтез широко распространенной в растениях рафинозы происходит путем ферментативного трансгликозилирования, при котором остаток галактозы переносится с уридиндифосфатгалактозы на сахарозу.
Синтез в растениях такого важного запасного углевода, как крахмал, происходит под действием фермента синтетазы крахмала при участин UDP глюкозы или АDРглюкозы.
В запасающих крахмал органах растений - клубнях картофеля, созревающих семенах риса и кукурузы - наиболее активный источник глюкозильных остатков – АDРглюкоза. Синтезированная амилоза дает затем амилопектин под действием энзима, который также является трансгликозилазой и получен в кристаллическом состоянии из клубней картофеля.
Биосинтез каллозы также происходит путем трансгликозилирования при участии UDPG как источника глюкозных остатков.
Реакция ферментативного трансгликозилирования позволяет объяснить известный факт чрезвычайно легкого превращения крахмала и инулина в сахарозу и образование этих сложных полисахаридов из сахарозы.
Известно, что крахмал, накапливающийся в листьях при фотосинтезе, может очень быстро превращаться в сахарозу - важнейшую транспортную форму углеводов в растении, в виде которой образовавшиеся при фотосинтезе углеводы перетекают из листа в семена, клубни и луковицы, где сахароза снова превращается в крахмал или инулин. Весьма существенно отметить при этом, что амилазы не принимают никакого участия в процессе превращения крахмала в сахарозу, поскольку мальтоза и декстрины не накапливаются в листьях и стеблях ассимилирующих растений.
В связи с важной ролью, которую играют инулин и левулёзаны в обмене веществ многих растений, заслуживают внимания факты обнаружения в растениях ферментов, катализирующих взаимные превращения полифруктозидов и сахарозы. Так, в молодых побегах земляной груши найден фермент, который в присутствии фосфатов катализирует превращение сахарозы в полифруктозиды и свободную глюкозу; из клубней артишока выделены ферментные препараты, катализирующие перенос остатков фруктозы с инулина на сахарозу и свободную фруктозу.
Приведенные данные свидетельствуют о чрезвычайной легкости, с которой происходят взаимные ферментативные превращения глюкозы, фруктозы, сахарозы, крахмала и левулёзанов (включая инулин).
На основании экспериментального материала можно наметить схему взаимных превращений этих углеводов в растениях. Центральное положение при этом занимают глюкозо-6-фосфат и фруктозо-6-фосфат, образующиеся в процессе фотосинтеза. Приведенная схема не дает представления о возможных путях образования пентоз в растении. Они могут образовываться при декарбоксилировании UDP уроновых кислот, чрезвычайно широко распространенных в растительных организмах в виде различного рода полиуронидов. Имеются экспериментальные данные, свидетельствующие о том, что ксилан синтезируется из ксилозы, которая образуется путем окисления глюкозы у шестого углеродного атома и последующего декарбоксилирования возникающей таким образом уроновой кислоты. Результаты этих опытов подтверждают представление о том, что декарбоксилирование галактуроновой и глюкуроновой кислот (или их полимеров) является важнейшим путем синтеза арабана и ксилана в растительном организме. Пентозы могут также возникать при декарбоксилировании кислот, образующихся при окислении молекулы гексозы у первого углеродного атома. Так, при декарбоксилировании фосфоглюконовой кислоты ферментными препаратами, выделенными из дрожжей, бактерий и высших растений, образуется фосфорный эфир кетопентозы - рибулозы; возникающий при этом рибулозофосфат под действием рибозофосфат-изомеразы дает рибозо-5-фосфат, превращающийся под влиянием фермента фосфорибомутазы в рибозо-1-фосфат.
Рибулоза под действием особой изомеразы может превращаться в арабинозу, а специфическая изомераза катализирует превращение рибулозо-5-фосфата в ксилулозо-5-фосфат. Таким образом, в результате ферментативных превращений фосфоглюконовой кислоты может возникать ряд пентоз и их фосфорных эфиров.
Наконец, образование пентоз можно представить как результат синтезирующего действия альдолазы. При взаимодействии фосфодиоксиацетона и фосфоглицеринового альдегида, происходящем под влиянием альдолазы, возникает фруктозодифосфат. Мейергофом показано, что под действием альдолазы фосфодиоксиацетон может обратимо конденсироваться не только с глицериновым альдегидом, но также с целым рядом других альдегидов, найденных в растениях, причем в результате этой реакции образуются пентозы. Таким образом, альдолаза, найденная во всех растениях, может катализировать биосинтез как гексоз, так и пентоз.
Несмотря на то, что пентозы могут образовываться тремя описанными путями, в настоящее время можно считать установленным, что у высших растений особенно важен путь, основанный на декарбоксилировании UDP уроновых кислот.
Уроновые кислоты - исходный материал для синтеза пектиновых веществ; это показано с помощью изотопной методики.
Во взаимопревращениях моносахаридов у растений, микроорганизмов и животных наряду с альдолазой важную роль играют ферменты транскетолаза и трансальдолаза. Таким образом, благодаря пентозофосфатному пути, альдолазе, транскетолазе и трансальдолазе происходят ферментативные взаимопревращения триоз, тетроз, пентоз, гексоз и гептоз.
Особенно глубокие превращения углеводов происходят при отложении их в запасных органах, подобных корневищам, клубням, луковицам и семенам, а также при прорастании, когда за счет веществ, отложенных в этих вместилищах запасов, происходит формирование тканей развивающегося молодого растения.
Процесс прорастания семян, клубней и луковиц сопровождается глубоким гидролизом отложенных в них высокомолекулярных полисахаридов, в первую очередь крахмала и инулина, с образованием растворимых углеводов. Благодаря резкому возрастанию активности a и b-амилаз происходит гидролиз крахмала с образованием декстринов и мальтозы; в клубнях и корнях, содержащих инулин, под действием инулазы происходит энергичный гидролиз инулина с образованием фруктозы и продуктов расщепления инулина - инулидов.
Уменьшение содержания крахмала сопровождается нарастанием количества декстринов, мальтозы и моносахаридов. Именно поэтому мальтоза, находящаяся в значительном количестве в прорастающем зерне, получила название солодового сахара. Накопление под действием амилазы значительных количеств декстринов определяет низкие хлебопекарные качества муки, полученной из проросшего зерна.
При прорастании семян, клубней и луковиц происходит не только гидролитическое расщепление крахмала и инулина с образованием соответствующих растворимых углеводов, но также накопление сахарозы благодаря действию трансгликозилирующих ферментов.
Прорастание семян сопровождается также глубоким гидролитическим расщеплением гемицеллюлоз, превращающихся под действием соответствующих ферментов (гемиuеллюлаз) в моносахариды. В течение длительного времени предполагали, что гемицеллюлозы, содержащиеся в очень больших количествах в некоторых семенах (например, у бобовых), не участвуют в обмене веществ. Однако исследования выдающегося швейцарского биохимика Эрнста Шульце показали, что гемицеллюлозы представляют собой весьма подвижную форму углеводов, мобилизуемую при прорастании семян и легко превращающуюся в сахара. Ферменты, катализирующие гидролиз гемиuеллюлоз, - гемицеллюлазы - найдены в прорастаюшем зерне, а также у некоторых бактерий и плесневых грибов.
Участие гемицеллюлоз в углеводном обмене растений показано также при изучении процессов во время созревания зерна. До недавнего времени предполагали, чтов созревающем зерне образование запасных углеводов - крахмала и гемицеллюлоз - происходит только за счет притекающих в зерно из листьев растворимых углеводов - сахаров и левулёзанов. Однако А. М. Палеевым установлено, что при созревании ржаного зерна отмечается также неуклонное уменьшение сухого вещества стебля и одновременное нарастание сухой массы колоса. Аналогичная картина наблюдается также у листьев, с той лишь разницей, что уменьшение абсолютно сухой массы листа начинается уже со времени колошения. По мере созревания колоса в листе и соломе снижается абсолютное содержание клетчатки, гемицеллюлоз и лигнина, используемых на построение крахмала и гемицеллюлоз зерна. Этот процесс Палеев называет раздревеснением. Таким образом, накопление крахмала и гемицеллюлоз в созревающем зерне идет не только за счет сахаров, образующихся в листьях при фотосинтезе, но и за счет растворимых углеводов, образующихся из клетчатки и гемицеллюлоз, которые содержатся в клеточных стенках листа и соломы.
Аналогичный процесс «перекачки» органических веществ из стеблей и листьев в клубни происходит на последних стадиях развития картофеля - накопление в клубнях крахмала и белка сопровождается уменьшением абсолютного количества органических веществ в ботве.
Возникает вопрос - каковы ферментативные реакции, лежащие в основе биосинтеза полисахаридов, образующих клеточные стенки растений, - клетчатки, хитина, различных гемицеллюлоз, а также пектиновых веществ.
В настоящее время можно считать установленным, что биосинтез этих полисахаридов происходит путем реакций перегликозилирования при участии определенных нуклеозидфосфатов. Так, в отношении клетчатки с полной определенностью установлено, что синтез ее идет непосредственно из глюкозы.
Биосинтез гемицеллюлоз и пектиновых веществ в растениях осуществляется также при участии различных ферментных систем, коферментами которых являются нуклеозиддифосфаты, в частности уридиндифосфат, а исходным материалом - D-глюкуроновая и D-галактуроновая кислоты.
Таким образом, биосинтез различных олигосахаридов и полисахаридов -сахарозы, трегалозы, рафинозы, крахмала, маннана, хитина, декстрана. левулезанов, гемицеллюлоз, пектина и клетчатки - происходит в результате ферментативных реакций трансгликозилирования, при этом роль коферментов и источников остатков того или иного сахара играют различные нуклеозиддифосфатсахара.
НАКОПЛЕНИЕ УГЛЕВОДОВ. В каждом вегетационном периоде накопление резервов питательных веществ необходимо для выживания многолетних растений тех мест, где зимы слишком холодны для фотосинтеза. Сказанное верно, в частности, в отношении лиственных деревьев, у которых поддержание жизни зимой и возобновление роста весной зависят от наличия питательных веществ, накопленных за предшествующий вегетационный период. Хотя главным запасным продуктом в древесных растениях является крахмал, но запасаются и другие вещества, в том числе жиры, азотистые соединения, сахароза, рафиноза, фруктозаны и гемицеллюлозы.
Баланс углеводов древесного растения можно выразить так:
Приход = углеводы, образовавшиеся при фотосинтезе.
Расход = углеводы, использованные на ассимиляцию и дыхание.
Сальдо = углеводы, накопленные-растением.
РАСПРЕДЕЛЕНИЕ УГЛЕВОДОВ. Запасные углеводы накапливаются преимущественно в клетках паренхимы. Гибель клеток сопровождается утилизацией или потерей запасов. Количество углеводов в разных частях древесных растений заметно различается. Наблюдаются также сезонные колебания количества и форм углеводов. Имеются также различия в характере сезонных колебаний накопления углеводов между листопадными и вечнозелеными видами тропиков и умеренной зоны.
Необходимо различать общее содержание и концентрацию углеводов в разных частях растений. Распределение углеводов часто выражают в процентах от сухого веса различных тканей. Это может ввести в заблуждение, так как высокие концентрации углеводов часто наблюдаются в тканях, составляющих небольшую долю сухого веса всего растения. Например, в молодых и старых деревьях концентрация углеводов в корнях обычно выше, чем в надземных частях. Тем не менее у взрослых деревьев именно надземные части являются основными вместилищами углеводов, поскольку ствол, ветви и листья имеют больший сухой вес чем корни.
У изученных Марником (1933, 1942) яблонь концентрация углеводов в корнях была выше, чем в стволах, но тем не менее надземные части, которые приблизительно в три раза тяжелее корней, содержали больше углеводов. Расделение углеводов в надземных и подземных частях также является функцией возраста растений, поскольку отношение масс подземной и надземной частей с возрастом постепенно уменьшается. В однолетних яблонях запасы углеводов распределялись между подземными и надземными частями почти поровну.
В листьях концентрация углеводов обычно высокая, но они составляют относительно небольшую долю всех углеводов древесного растения. Хвоя сосны виргинской, исследованной Гептингом (1945), содержала 16,6% углеводов, что почти в 7 раз превышает их концентрацию в древесине ствола. Содержание углеводов (крахмал плюс сахара) в листьях яблони доходило до 9%, что намного больше величин, приводимых для других тканей. Общее содержание углеводов в листьях составляло лишь примерно 5% запасов углеводов всего дерева.
ИСПОЛЬЗОВАНИЕ ЗАПАСНЫХ УГЛЕВОДОВ. Наиболее очевидно использование запасных углеводов для поддержания дыхания и роста в периоды, когда питательные вещества не поступают непосредственно от фотосинтеза. Вся метаболическая активность в ночное время зависит от питательных веществ, накопленных за день. У листопадных деревьев дыхание и другие процессы поддерживаются зимой благодаря использованию запасов питательных веществ. По исследованиям, у молодых яблонь менее четверти углеводного запаса использовалось весной на рост новых тканей, а большая часть оставшихся углеводов была использована на дыхание.
Запасные углеводы, помимо их роли в поддержании дыхания в ночное время и зимой, важны и в начале вегетационного периода, что подчеркивается фактом роста корней многих видов листопадных деревьев до распускания листьев. Образование поросли и корневых отпрысков также зависит от мобилизации углеводных запасов.
Клэрк и Лиминг (1953) обнаружили, что отрастание дуба после кольцевания происходило в первый и второй сезоны, пока не исчерпались запасы углеводов. У деревьев, окольцованных на более позднем этапе вегетационного периода, отрастание в следующем году замедлялось. По наблюдениям Тью (1970), продолжительность периода, в течение которого у осины образовывались корневые отпрыски, зависела от количества запасных углеводов. Шир и Засада (1973) нашли положительную корреляцию между массой корневых отпрысков осины и количеством запасных углеводов. Эти наблюдения показывают, что отрастание уменьшается по мере исчерпания запаса углеводов. С другой стороны, Венджер (1953) сообщил о втором минимуме отрастания у амбрового дерева в конце лета, когда содержание углеводов в корнях было относительно высоким. Он сделал вывод о том, что регуляторы роста играли более важную роль, чем запасы питательных веществ.
При рубке деревьев в начале лета, когда их листва только что распустилась и запасы углеводов малы, пневая поросль развивается слабо по сравнению с обильным отрастанием пней деревьев, срубленных в период покоя.
Вследствие отсутствия у лиственных деревьев умеренной зоны фотосинтезирующих листьев во время набухания почек и увеличения их сухого веса начальные этапы роста побегов зависят от углеводных запасов.
Случай особой важности запасных углеводов для роста побегов отметил Шимпер (1903), сообщавший, что почки тропического вида Brownia несколько дней разрастались со скоростью 2,6 см в день, в то время как листья оставались свернутыми и фотосинтеза не было. Следовательно, рост побегов поддерживался за счет запасов углеводов, передвигавшихся из других частей дерева. Другой пример значения запасов демонстрирует появление второй партии листвы из покоящихся почек у деревьев умеренной зоны весной, после гибели первой партии молодых листьев от заморозков.
Важность запасов для листопадных деревьев еще раз подчеркивают: во-первых, быстрое исчезновение запасных углеводов после распускания почек и развития побегов, и, во-вторых, мобилизация меченых радиоактивным углеродом запасов. Согласно подсчетам Хенсена (1971), от 1/2 до 2/3 углеводов, необходимых для роста цветков и побегов яблони в самом начале вегетационного периода, поступали скорее из запасов, чем от продуктов текущего фотосинтеза. Это относилось только к периоду появления 5-6 первых листьев, после которого рост плодов и побегов был связан с утилизацией непосредственных продуктов фотосинтеза.
В отличие от листопадных вечнозеленые деревья к началу ежегодного роста побегов уже располагают функционирующей фотосинтетической системой. Например, у сосен синтезированные старой хвоей углеводы используются уже на самых ранних этапах удлинения, побегов. Как и многие листопадные деревья, вечнозеленые не переносят потери кроны. Такие наблюдения привели к сомнениям относительно значения запасных углеводов для роста побегов у вечнозеленых. Однако имеются веские доказательства того, что запасные углеводы все-таки играют важную роль в росте вечнозеленых растений. Так, в рано начинающемся росте корней утилизируется значительная доля запасов углеводов. Кроме того, накопление и использование запасов - важная особенность роста надземных тканей.
Часть ассимилятов, образующихся в вечнозеленых деревьях умеренной зоны в конце сезона, сохраняется и используется на рост в следующем вегетационном сезоне. Весной запасенные в ветвях углеводы использовались на рост побегов. Использование запасов на рост побегов у голосеменных подтверждают наблюдения за быстрым исчезновением углеводов из ветвей при развитии побегов.
Например, от апреля до начала июня, когда новые побеги дугласии бурно развивались, количество углеводов в однолетних побегах быстро уменьшалось. Концентрация и количество углеводов в новых, развивающихся побегах увеличивались от раскрытия почек до середины июня. У молодых растений тиса большая часть углеводов, использованных на рост побегов, хранилась до утилизации в старых листьях.
СЕЗОННАЯ ЦИКЛИЧНОСТЬ СОДЕРЖАНИЯ УГЛЕВОДОВ. Древесные растения накапливают запасы углеводов в периоды их избыточного образования, а затем расходуют их, когда скорость утилизации превышает скорость новообразования. В частности, во время наиболее быстрого роста запасные углеводы исчезают из ветвей, стволов и корней в порядке, зависящем от видовых особенностей роста. Сезонные циклы содержания углеводов наиболее выражены у многих листопадных деревьев умеренной зоны. Общее содержание углеводов в стволах и ветвях достигает максимума осенью, незадолго до начала листопада. В конце зимы оно начинает уменьшаться и быстро уменьшается ранней весной, когда углеводы используются на усиленное дыхание и рост новых тканей.
У многих видов обнаружены два сезонных максимума содержания крахмала: весной и в конце лета или начале осени. Вслед за весенним максимумом содержание крахмала падает в результате утилизации резервов на рост побегов и камбия. Содержание крахмала зимой снижается вследствие его превращения при низких температурах в сахарозу.
Годичные циклы углеводов у видов с периодическим ростом характеризуются исчезновением углеводов с каждой вспышкой роста, после чего новые углеводы занимают их место. Годичные циклы углеводов у вечнозеленых деревьев отличаются от таковых у листопадных деревьев. Вечнозеленые виды накапливают углеводы зимой намного позже. Сезонные колебания запасов углеводов у них намного меньше, чем у листопадных деревьев.
Так, в ветвях каштана калифорнийского, у которого листва опадает при засухе, содержание углеводов в ветвях в течение года варьировало гораздо больше, чем у вечнозеленого дуба траволистного. У каштана калифорнийского осенью, в период образования плодов, стебли были без листьев, а содержание углеводов снижалось с 17 до 10%. Когда в феврале развивались новые листья, запасы углеводов расходовались еще быстрее, пока не уменьшались до самого низкого в сезоне уровня - 5%. В марте и апреле, когда деревья были полностью покрыты листвой, содержание углеводов в ветвях восстанавливалось до самых больших за год значений. По сравнению с этим содержание углеводов в облиственных стеблях дуба траволистного было постоянным, варьируя приблизительно от 3,5 до 5%.
Даже у деревьев, растущих в тропическом климате, отмечаются годичные циклы содержания углеводов, хотя в общем диапазон этих циклов ограничен по сравнению с такими же циклами у деревьев более холодных климатов. У периодически растущих тропических деревьев с каждым усилением роста наблюдается некоторое истощение углеводов.
ОСЕННЯЯ ОКРАСКА. Антоциановые пигменты, вызывающие розовую, красную и пурпурную осеннюю окраску листьев, связаны с углеводами и накопление углеводов способствует их образованию. Антоцианы — это гликозиды, возникающие при соединении различных сахаров с циклическими соединениями, называемыми антоцианидинами. Антоцианы растворимы в воде и обычно встречаются в клеточном соке. В кислых растворах они имеют красный цвет, а с увеличением рН могут становиться фиолетово-синими. Количество антоциановых пигментов зависит прежде всего от наличия определенных наследственных факторов их образования, но окружающая среда также оказывает влияние.
С понижением температуры осенью листья перестают образовывать хлорофилл. В это время некотрые виды, содержащие много углеводов и обладающие наследственной способностью, начинают образовывать в листьях антоцианы. После прекращения синтеза хлорофилла имеющийся хлорофилл начинает разрушаться, и только что образовавшиеся антоцианы становятся заметными. У деревьев, не образующих антоциановых пигментов, осенний распад хлорофилла обнаруживает относительно более стабильные желто-оранжевые каротины и ксантофиллы, что придает листьям светло-желтую окраску, или же примесь красного антоциана в желтом каротине, что дает ярко-оранжевый цвет (как у некоторых видов клена). У других видов хлорофилл и каротиноиды распадаются одновременно, и синтезируются новые каротиноиды. Так путем разрушения зеленых пигментов, выявления желтых, образования красных пигментов или всеми тремя способами сразу листья могут приобретать различные оттенки желтого, оранжевого, малинового, пурпурного и красного цветов.
У таких деревьев, как ольха и белая акация, изменения окраски выражены слабо. В противоположность им, листья большой группы растений, включающей орех черный, катальпу, вяз, карию, липу и платан становятся ржаво-зелеными и желтыми. Окраска листьев тополей, гледичии трехколючковой, гинкго, бука и большинства видов березы изменяется в различные оттенки желтого цвета. Но, несомненно, наибольшее впечатление оставляют красные тона клена красного и сахарного, сассафраса лекарственного, сумаха, дуба белого и американского шарлахового, ирги и бересклета крылатого - растений, образующих большие количества антоциановых пигментов. Разные виды и экземпляры клена демонстрируют постепенный переход окраски от желтой до темно-красной.
Деревья одного и того же вида, растущие вместе, часто сильно различаются по окраске благодаря индивидуальным различиям в количестве растворимых углеводов. Некоторые окрашиваются позже других. Дубы окаршиваются поздней осенью, обычно после того, как наиболее выраженная окраска кленов успевает поблекнуть. Желто-коричневая окраска бука и некоторых видов дуба вызывается присутствием в листьях одновременно и желтых каротиноидов и танинов.
Различия в скорости изменения осенней окраски у разных видов свидетельствуют о глубоких различиях интенсивности распада хлорофилла. По сообщению Вицковского (1958), в то время как у одного из видов магнолии наблюдалось быстрое разрушение хлорофилла (за 35 дней), у шелковицы белой распад происходил медленно (более 60 дней). Перед опадением листья явора и бука теряли весь хлорофилл, тогда как листья сирени только 40%. Вольф (1956) показал широкое варьирование содержания хлорофилла в листьях и скорости его распада осенью. У многих видов хлорофилл а разрушался быстрее хлорофилла b.
Гудвин (1958) проследил изменения содержания хлорофилла и каротиноидных пигментов с июня по ноябрь в сливе черной, дубе черешчатом и яворе. У дуба и явора содержание хлорофилла и каротиноидов уменьшалось почти до нуля. У дуба они расходовались одновременно, тогда как у явора падение содержания хлорофилла предшествовало снижению содержания каротиноидов. У сливы каротиноиды первыми проявляли тенденцию к уменьшению, но содержание их и хлорофилла снижалось только наполовину. По другим данным, при изменении окраски листьям клена образовывался каротиноидный пигмент, отличавшийся от имевшихся летом каротиноидов, а общее содержание каротиноидов уменьшалось.
Любой фактор, влияющий на синтез углеводов или на превращение нерастворимых углеводов в растворимые, способствует образованию антоциана и появлению ярких осенних окрасок. К наиболее важным факторам окружающей среды, определяющим осеннюю окраску, относятся температура, свет и водоснабжение. Понижение температуры ниже нуля способствует образованию антоциана. Ранние суровые морозы делают красные осенние цвета менее яркими, чем они были бы без них. Яркий свет также способствует появлению красной окраски, так как антоциановые пигменты обычно образуются в листьях, находящихся на свету. Если во время образования красных пигментов один лист был накрыт другим, в нижнем листе красный пигмент обычно не образуется. Снабжение водой также влияет на образование антоцианов: засуха способствует появлению ярко-красной окраски. Дождливые дни с недостатком света незадолго до периода наивысшего развития окраски существенно уменьшают яркость красок листопада. Следовательно, самые лучшие осенние цвета наблюдаются при ясной, сухой и прохладной (но не морозной) погоде.
Дата добавления: 2015-02-03; просмотров: 4852;