Синтез и распад жиров
Энергичное образование жиров за счет углеводов происходит в созревающих семенах и плодах, накапливающих значительное количество жиров.
Так же как и у микроорганизмов, жир из углеводов в созревающих семенах и плодах растений образуется при достаточном доступе кислорода, так как часть потребляемого сахара окисляется полностью до диоксида углерода и воды, а выделяемая при этом энергия используется на процесс синтеза жира. Вместе с тем образующиеся из сахара жирные кислоты содержат значительно меньше кислорода (около 11- 12%), чем исходный сахар, например глюкоза (около 50%). Поэтому кислород, необходимый для осуществления синтеза жирных кислот, частично берется из самого сахара, и потребление атмосферного кислорода снижается. В результате дыхательные коэффициенты у созревающих масличных семян, так же как и у микроорганизмов, образующих жир из сахара, значительно превышают 1 (дыхательные коэффициенты у созревающих семян клещевины достигают 4,71).
Составные части жира - глицерин и жирные кислоты - образуются из сахаров. Главным источником компонентов жира являются гексозы, в первую очередь глюкоза и фруктоза. Однако исходным веществом, используемым на синтез жира в растительном организме, могут быть не только гексозы, но также пентозы и продукты глубокой диссимиляции углеводов, содержащие два или три углеродных атома в молекуле. Глицерин, необходимый для синтеза жиров, образуется в процессе анаэробной диссимиляции углеводов путем восстановления глицеринового альдегида, получающегося из фруктозодифосфата под действием фермента альдолазы. Исходным материалом для синтеза насыщенных жирных кислот служит активный ацетил в виде ацетилкофермента А. Для этого процесса необходимы Мn2+ и СО2, который вступает в реакцию с ацетилкоферментом А, образуя малонилкофермент А - важнейший промежуточный продукт ферментативного синтеза жирных кислот. В процессе присоединения CO2 к ацетилкоферменту А важную каталитическую роль играет биотин, а источником энергии служит АТР. Схематически процесс биосинтеза малонилкофермента А можно представить следующим образом:

Таким образом, фактический источник двууглеродного фрагмента, последовательно присоединяющегося при синтезе жирных кислот, - малонилкофермент А. Биотин же является коферментом фермента ацетил-СоА - карбоксилазы, который в процессе синтеза жирных кислот катализирует реакцию присоединения СО2.
Высшие насыщенные жирные кислоты превращаются в ненасыщенные в результате реакций, в которых участвуют NADPH и кислород. Например, олеиновая кислота может синтезироваться из стеариновой кислоты путем отнятия отпоследней водорода. В листьях синтез жирных кислот осуществляется не в хлоропластах, а в семенах - в пропластидах. В этих органеллах синтезируется пальмитиновая кислота, которая затем, при участии кислорода и ферредоксина, окисляется и превращается в олеиновую кислоту. Эта последняя, уже вне хлоропластов или пропластид, подвергается разнообразным модификациям - гидроксилированию, восстановлению, конденсации. В результате образуется все многообразие содержащихся в растениях жирных кислот.
Процесс расщепления жира в растительном организме происходит особенно энергично при прорастании масличных семян. Он начинается с гидролитического распада жиров, происходящего под действием липазы и сопровождающегося накоплением глицерина и свободных жирных кислот. Образующиеся глицерин и жирные кислоты чрезвычайно быстро используются для различных синтезов, протекающих в развивающемся ростке. При этом главным продуктом, возникающим в результате превращения жиров, является сахар. Необходимо отметить, что при прорастании богатых жиром семян образуются не только гексозы, но и пентозы. Этот факт указывает на то, что во время прорастания семени жир расщепляется до низкомолекулярных соединений, содержащих два или три углеродных атома в молекуле. Путем конденсации этих низкомолекулярных соединений образуются затем различные моносахариды и другие вещества.
Важнейший этап диссимиляции жирных кислот - b -окисление, когда окисление жирной кислоты происходит у того углеродного атома, который находится в b-положении по отношению к карбоксильной группе.
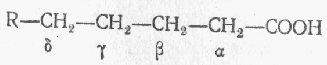
В результате происходит разрыв углеродной цепочки жирной кислоты между a- и b-углеродными атомами с образованием ацетильного радикала и новой высокомолекулярной жирной кислоты, содержащей на два углеродных атома меньше, чем подвергшаяся окислению первоначальная жирная кислота. Процесс b-окисления жирных кислот осуществляется при участии кофермента А и начинается с его присоединения к молекуле жирной кислоты.
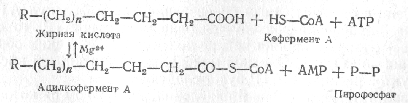
Затем происходит отнятие водорода в a-b-положении, осуществляемое под действием флавинового фермента. Далее по месту двойной связи присоединяется молекула воды и образует b-оксикислота. Образовавшаяся b-оксикислота подвергается окислению путем отнятия водорода, которое происходит при участии NAD+, что приводит к образованию b-кетокислоты.
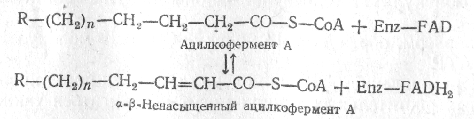
Последний этап b-окисления жирной кислоты - расщепление возникающей b-кетокислоты, происходящее под действием новой молекулы кофермента A.
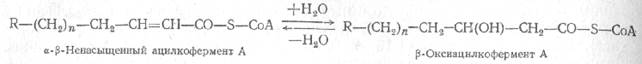
В результате образуется ацетилкофермент А и соединенный с другим остатком коэнзима А радикал новой жирной кислоты, содержащей на два углеродных атома меньше, чем молекула исходной жирной кислоты.
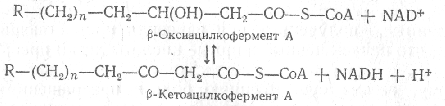
Новая кислота может снова подвергнуться b-окислению, пока не окислится полностью.
Таким образом, конечный продукт b-окисления жирных кислот -ацетилСоА. Он либо полностью окисляется до CO2 и H2O, вступая в цикл трикарбоновых кислот, либо используется на синтез углеводов в глиоксилатном цикле. FADH2 и NADH, образующиеся при b-окислении, окисляются в дыхательной цепи с одновременным образованием пяти молекул АТР. Так как на первом этапе b-окисления расходуется одна молекула АТР, то «чистый» выход АТР на первом этапе составляет четыре молекулы АТР. Таким образом, при полном окислении до СО2 и H2O одной молекулы пальмитиновой кислоты C16H32О2 выход продуктов реакции и АТР будет следующий.
1. В результате семи этапов b-окисления, на каждом из которых образуется пять молекул АТР, возникает 35 молекул АТР. С вычетом одной молекулы АТР, расходуемой на первом этапе, получаем 34 молекулы.
2. Полное окисление восьми молекул ацетил-СоА в цикле трикарбоновых кислот дает 96 молекул АТР (окисление одной молекулы ацетил-СоА дает 12 молекул АТР).
3. Таким образом, образуется 16 молекул СО2 и 16 молекул Н2О и возникает 130 молекул АТР.
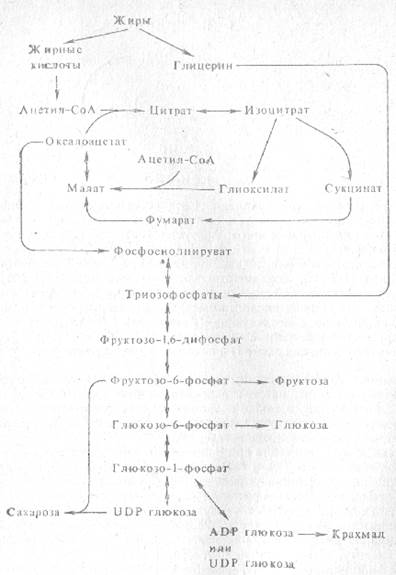
Ацетил-СоА, возникающий при р-окислении жирных кислот, может не только полностью окисляться до СО2 и Н2О, но также использоваться на синтез различных соединений, в частности углеводов. Именно этот процесс синтеза углеводов из жира происходит при прорастании богатых жиром семян. При этом ацетил-СоА, образующийся в результате b-окисления, включается в реакции цикла глиоксилевой кислоты и дает щавелевоуксусную кислоту, а затем яблочную кислоту, которая далее превращается в фосфоенолпировиноградную кислоту, а последняя - в углеводы.
Ранее было указано, что превращение жиров в сахара при прорастании богатых маслом семян происходит в особых субклеточных структурах - глиоксисомах. Этот процесс получил название гликонеогенеза. Жир под действием липаз гидролизуется до жирных кислот и глицерина. Жирные кислоты, окисляясь, образуют ацетил-СоА, который через серию реакций глиоксилатного цикла, локализованных главным образом в глиоксисомах, дает щавелевоуксусную кислоту, превращающуюся далее в фосфоенолпируват. Глицерин же дает фосфотриозы, которые, как и фосфоенолпируват, служат исходным материалом для синтеза глюкозы и фруктозы путем обращения процесса анаэробного расщепления глюкозы.
Наличие ферментативных реакций, лежащих в основе такого превращения, доказано для прорастающих семян клещевины, тыквы, арахиса и подсолнечника.
Дата добавления: 2015-02-03; просмотров: 4005;