Филогения типа Spongia
В организации губок много признаков большой примитивности: отсутствие настоящих дифференцированных тканей и органов, чрезвычайная пластичность клеточных элементов, отсутствие резко выраженной индивидуальности в колониях — все это свидетельство того, что губки— просто устроенные представители многоклеточных.
Если принять теорию Мечникова о происхождении многоклеточных (с. 93), то легко видеть, что личинка, свойственная большинству губок,— паренхимула (см. рис. 86), по строению почти полностью соответствует гипотетической мечниковской фагоцителле. У нее имеется поверхностный, эктодермальный слой жгутиковых клеток и внутренний рыхлый слой клеток — энтодерма. Можно предположить, что фагоцителла перешла к сидячему образу жизни и таким путем дала качало типу губок. При этом, как уже отмечалось (с. 109), судьба клеточных слоев фагоци-теллы у губок оказалась иной, чем у прочих многоклеточных («извращение» зародышевых листков): наружный эктодермальный слой жгутиковых клеток у губок дал начало пищеварительному слою хоаноцитов, который вместе с тем осуществляет кинетическую мерцательную водо-движущую функцию; внутренние энтодермальные клетки зародыша, которые у других групп животных дают начало энтодермальной кишке, у губок превращаются в клетки поверхности тела (дермальные) и в клеточные элементы мезоглеи. Все эти факты говорят о том, что отделение губок от ствола многоклеточных произошло очень рано, еще до того, как определилась окончательная судьба двух основных клеточных пластов тела. Некоторые зоологи считают, что губки произошли от колониальных воротничковых жгутиконосцев независимо от прочих многоклеточных. Другие полагают, что многоклеточные происходят общим стволом, от которого очень рано отделились губки. Второй взгляд представляется более обоснованным потому, что личинка — паренхимула губок — сходна с планулой кишечнополостных. Это говорит об общности их происхождения.
Губки — очень древние организмы. Их ископаемые остатки многочисленны в кембрийских морских отложениях. Встречаются они и в протерозойских породах.
НАДРАЗДЕЛ EUMETAZOA
РАЗДЕЛ ЛУЧИСТЫЕ (RADIATA)
ТИП КИШЕЧНОПОЛОСТНЫЕ (COELENTERATA, ИЛИСМОАША)
Кишечнополостные ведут исключительно водный и в большинстве случаев морской образ жизни. Одни из них свободно плавают, другие, не менее многочисленные формы — сидячие и прикрепленные ко дну животные. К Coelenterata относится около 9000 видов.
Строение кишечнополостных характеризуется радиальной, или лучистой, симметрией. В теле их можно различить одну главную продольную ось, вокруг которой в радиальном (лучистом) порядке расположены различные органы. От числа повторяющихся органов зависит порядок радиальной симметрии. Так, если вокруг продольной оси располагается 4 одинаковых органа, то радиальная симметрия в этом случае называется четырехлучевой. Если таких органов шесть, то и порядок симметрии будет шестилучевым, и т. д. Ввиду подобного расположения органов через тело кишечнополостных можно всегда провести несколько (2, 4, 6, 8 и более) плоскостей симметрии, т. е. плоскостей, которыми тело делится ка две половины, зеркально отображающие одна другую. В этом отношении кишечнополостные резко отличаются от двустороннесимметрич-ных, или билатеральных, животных (Bilateria), у которых всего одна плоскость симметрии, делящая тело на две зеркально подобные половины: правую и левую.
Радиальная симметрия встречается у нескольких далеко друг от друга стоящих групп животных, которые, однако, имеют общую биологическую черту. Все они или ведут в настоящее время сидячий образ жизни, или вели его в прошлом, т. е. происходят от прикрепленных животных. Отсюда можно сделать вывод, что сидячий образ жизни способствует развитию лучистой симметрии.
Биологически это правило объясняется тем, что у сидячих животных один полюс служит обычно для прикрепления, другой, свободный, несет на себе рот. Свободный ротовой полюс животного по отношению к окружающим предметам (в смысле возможности захвата пищи, осязания и т. п.) поставлен со всех сторон в совершенно одинаковые условия, вследствие чего многие органы и получают одинаковое развитие на разных пунктах тела, расположенных вокруг главной оси, проходящей через рот до противоположного ему прикрепленного полюса; результатом этого является выработка лучистой симметрии. Совсем иначе дело обстоит у ползающих животных.
Кншечнополостные—-двухслойные животные (Diploblastica): в онтогенезе у них формируются-только два зародышевых листка — экто- и
Н4

|

|
|

|
| ~*т А |
| В |

Рис. 90. Старинные изображения различных Eumetazoa. А — пресноподный малоще-тинковый червь в изображении Розель фон Розснгофа (1775); Б — водяная блоха — дафния (из Франсэ), фантастически нарисована голова с птичьим клювом, глазом и бровью; В — ракообразные, слева — изображение краба на древней греческой монете, форма тела и конечности животного переданы необычайно правдиво; справа — изображение морского таракана (рачок из отр. Isopoda по Себастиану Мюнстсру, 1550), рисунок обнаруживает полное незнание автором изображаемого животного; Г — древнеегипетский рисунок жука-скарабея, следует отметить совершенно неправильное изображение «крыльев», напоминающих крылья птиц; Д — изображение осьминога на критской вазе (около 1500 лет до н. э.), рисунок поражает живостью передачи и значительной точностью, если не считать раздвоенности конца тела; Е— морские звезды (Олаф Магнус, середина XVI в.), интересен антропоморфизм рисунка (человеческие глаза, нос и рот) и совершенно неправильная ориентировка животных
(рот кверху, а не книзу)
энтодерма, отчетливо выраженные и у взрослого животного. Эктодерма и энтодерма разделены прослойкой мезоглеи.
В наиболее простом случае тело кишечнополостных имеет вид открытого на одном конце мешка. В полости мешка, выстланной энтодермой, происходит переваривание пищи, а отверстие служит ртом. Последний обычно окружен несколькими или одним венчиком щупалец, захватывающих пищу. Непереваренные остатки пищи удаляются из тела через ротовое отверстие. По строению наиболее просто организованные из кишечнополостных могут быть сведены к типичной гзструле.
В зависимости от образа жизни эта схема строения может несколько изменяться. Наиболее близки к ней сидячие формы, которым дано общее наименование — полипы: свободноплавающие кишечнополостные испытывают обычно сильное уплощение тела по направлению главной оси — это медузы. Деление на полипов и медуз не систематическое, а чисто морфологическое; иногда один и тот же вид кишечнополостных на различных стадиях жизненного цикла имеет строение то полипа, то медузы. В медузоидном состоянии кишечнополостные, как правило, одиночные животные. Напротив, полипы лишь в редких случаях'\5ывают одиночными. Громадное большинство их, начиная жизнь как одиночный полип, образует затем посредством почкования, не доходящего до конца, колонии, состоящие из сотен и тысяч особей. Колонии состоят из вполне одинаковых особей (мономорфные колонии) или же из особей, имеющих различное строение и выполняющих различные функции (полиморфные колонии).
Характернейшая черта типа — наличие стрекательных клеток. Движение осуществляется путем мускульных сокращений. Тип распадается па классы: Hydrozoa (гидрозои); Scyphpzoa (сцифоидные медузы); Anthozoa (коралловые полипы).
КЛАСС I. ГИДРОЗОИ (HYDROZOA)
Низший класс, состоящий большей частью из мелких форм, содержит полипов и медуз (2700 видов). В отличие от сцифомедуз и коралловых полипов полипы и медузы, принадлежащие к Hydrozoa, называются гидроидными.
ПОДКЛАСС I. ГИДРОИДНЫЕ (HYDROIDEA)
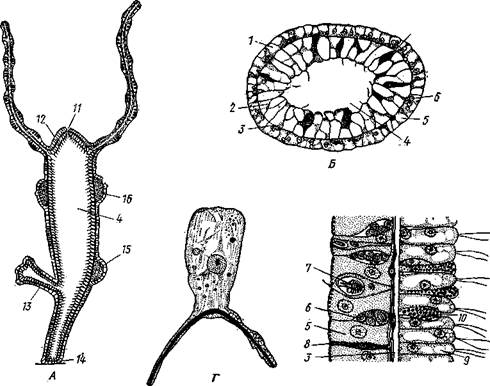
Строение гидры (Hydra).На примере гидры можно ознакомиться со строением гидроидных полипов. Гидра — одни из наиболее просто устроенных полипов. Это маленький (около 1 см) пресноводный полип, часто встречающийся в озерах и прудах. Тело гидры в виде продолговатого мешочка, прикрепляется к субстрату своим основанием, или подошвой; па свободном конце тела па особом, возвышении — ротовом конусе лежит рот, окруженный венчиком из 6—12 щупалец (рис. 91, А). Вся поверхность тела, вплоть до краев ротового отверстия, покрыта эктодермой, состоящей из нескольких сортов клеток. Большая часть ее образована цилиндрическими или кубическими эпителиальными клетками, основание которых, обращенное к мезоглее, вытягивается по направлению кверху и книзу (по продольной оси животного) в длинный отросток, лежащий параллельно поверхности тела (рис. 91, Г). Цитоплазма отростка дифференцируется в виде тончайших сократительных волоконец; отросток имеет значение мускульного. Цилиндрическая часть клетки входит в состав покровного однослойного эпителия. Эти клетки называются эпителиально-мускульными. Совокупность отростков всех таких
| I |
клеток образует в основании эпителия слой мускульных образований, совпадающих с продольной осью тела. При их одновременном сокращении тело полипа сильно укорачивается.
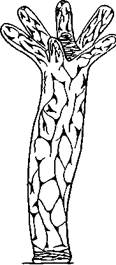
Между основаниями более крупных эпителиально-мускульных клеток располагаются мелкие промежуточные (интерстициальные) клетки. За их счет формируются половые и стрекательные клетки. Непосредственно под эпителием рассеяны нервные клетки звездчатой формы, которые своими отростками сообщаются между собой и образуют субэпителиальное нервное сплетение. Таким образом, нервная система гидры стоит на самой низкой ступени развития, имеет рассеянный, диффузный характер (рис. 92). Впрочем, даже у гидры отмечаются два сгущения нервного сплетения — вокруг рта и на подошве.
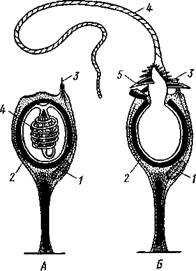
Характерная черта кишсчнополостных — присутствие в покровах стрекательных клеток (рис. 93, А, Б). Они развиваются из промежуточных клеток и содержат особую овальную стрекательную капсулу с плотными стенками. Капсула наполнена жидкостью, а на одном конце капсулы стенка ее впячена внутрь в виде очень тонкого, но полого отростка, который закручивается в капсуле в спирально завитую стрекательную нить. Стрекательные клетки служат гидре орудием нападения и защиты.
|
| •ю- |
| •В |
Рис. 91. Гидра Hydra oligactis. A—продольный разрез; Б — поперечный разрез; В — участок среза при большом увеличении; Г — эктодермальная эпителиально-мускульная
клетка (А — из Бриана, Б — по Полянскому, В — по Кестнеру, Г — по Роскину): / — эктодерма, 2 — энтодерма, 3 — базальная мембрана, 4 — гастральная полость, 5 —эктодер-мальные эпителиально-мускульные клетки, 6 — интерстициальные клетки, 7 — стрекательные клетки, 8 — нервные клетки, 9 — энтодермальные " эпителиально-мускульные клетки, 10 — железистые клетки, // — ротовое отверстие, 12 — ротовой конус, 13 — почка, 14 — подошва, 15 — яйцеклетка, /6 —
мужские гонады
. 117
|
| Рис. 92. Схема расположения нервных клеток в теле гидры (по Гессе) |
На наружной поверхности клетки имеется тонкий чувствительный волосок— книдоциль. Изучение стрекательных клеток с помощью электронного микроскопа показало значительную сложность строения книдоциля (рис. 93, В). Он состоит из длинного жгутика, окруженного 18—22 тонкими пальцевидными выростами цитоплазмы — микроворсинками. По строению жгутик книдоциль очень сходен со жгутиками и ресничками простейших, но в отличие от них неподвижен. При прикосновении добычи или врага к жгутику последний отклоняется и задевает одну или несколько микроворсинок, что приводит к возбуждению стрекательной клетки. При этом стрекательная капсула выбрасывает выворачивающуюся из нее наружу упругую нить, которая распрямляется, как стрела. Нить наподобие гарпуна усажена обращенными назад шипиками, а в основании несет более крупные шипы. Уколы нити ядовиты и могут парализовать мелких животных. После выбрасывания нити стрекательная клетка погибает. У гидры имеется несколько категорий капсул, функции которых различны. Рассмотренные крупные капсулы, служащие для пробивания покровов и поражения добычи, называются пенетрантами (рис. 93). Значительно более мелкие — вольвенты имеют короткие спирально закрученные нити, которые обвиваются вокруг различных выступов (щетинок, волосков и т. п.) на теле добычи и таким путем удерживают ее. Наконец, вытянутые стрекательные капсулы — глютинанты — приклеиваются к телу добычи длинными липкими нитями.
|
Энтодерма выстилает всю гастраль-ную (пищеварительную) полость вплоть до краев рта. В состав энтодермы входит также несколько кате-
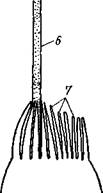
Рис. 93. Стрекательные клетки. А — в покоящемся состоянии;
Б — с выброшенной стрекательной нитью (по Кюну); В —
строение книдоциля (по Слаутербаку):
Дата добавления: 2015-01-24; просмотров: 1760;