Филогения типа Mollusca
Происхождение типа моллюсков и их филогенетические связи с другими группами беспозвоночных животных были и остаются предметом оживленной дискуссии зоологов. Долгое время доминировала точка зрения, согласно которой моллюски в своем происхождении связаны с ан-нелидами. И действительно, в эмбриональном развитии моллюсков ряд признаков указывает на родство их с кольчатыми червями: спиральное детерминированное дробление, способ закладки мезодермы, личинка трохофора. В строении взрослых моллюсков сходны с кольчецами отношения между гонадами, целомом и целомодуктами.
Более того, самые примитивные моллюски Amphineura и Monopla-cophora обнаруживают ясно выраженную метамерию, остатки которой в виде двух пар ктенидиев, почек и предсердий сохраняются у низших головоногих (Nautilus). Однако не все проявления метамерности могут приниматься во внимание. Расчленение раковины у Loricata и упорядоченное расположение мускульных пучков, так же как и большое количество жабр, вряд ли должны рассматриваться как пример сохранения моллюсками исходной метамерности. По мнению многих зоологов, это вторичное явление. Так, из большого и варьирующего у разных видов числа жабр только одна пара, расположенная около почечного отверстия и снабженная осфрадиями, вероятно, соответствует настоящим кте-
кидиям; остальные жабры возникли вторично для интенсификации функции дыхания.
С аналогичным явлением мы сталкиваемся и у современных Monopla-cophora. Внешняя простота колпачковидной раковины и метамерное расположение мышц явно вторичны, так как наиболее древние ископаемые представители группы обладали спиральной раковиной и всего одной парой раковинных мышечных пучков.
Значительно больший вес имеют следы метамерии в строении внутренних органов и в первую очередь органов, связанных с целомом. Наличие у Neopilina 6—7 пар целомодуктов, выполняющих функции почек и частично служащих для выведения половых продуктов, двух пар гонад и двух пар предсердий позволяет предполагать, что тело моллюсков первоначально состояло из небольшого числа (6—7) сегментов, унаследованных от далеких предков. Последними, по-видимому, были какие-то древние олигомерные кольчецы, тело которых состояло только из лар-вальных сегментов. Таким образом, тело современных моллюсков соответствует ларвальному телу кольчецов и вторично утратило метамерное строение. Уменьшение числа ряда образований (целомодуктов, предсердий, настоящих ктенидиев и т. д.) должно рассматриваться как результат олигомеризации гомологичных органов.
Среди современных моллюсков наиболее примитивными, по-видимому, являются моноплакофоры.
К особенно древним чертам организации Neopilina, несомненно, относятся: метамерия, двураздельность сердца и наличие продольных нервных стволов.
По наличию двух желудочков сердца Monoplacophora сравнимы с зародышами пластинчатожаберных, брюхоногих и головоногих, у которых сердце сначала представлено парой зачатков, лежащих по бокам от кишки. При дальнейшем развитии зачатки сливаются, образуя один желудочек.
У некоторых низших представителей класса Lamellibrar.chia (Area) слияния зачатков также не происходит, и взрослое животное имеет два сердца (с. 478).
Таким образом, двураздельное сердце Monoplacophora представляет очень примитивную особенность, у большинства Conchifera проявляющуюся лишь в онтогенезе. У Amphineura непарное сердце.
Из сказанного ясно, что кл. Monoplacophora во многих отношениях примитивнее современных панцирных. Естественно поэтому сделать вывод, что оба ствола моллюсков — Amphineura и Conchifera разделились очень рано. Monoplacophora еще очень близки к общему предку всех моллюсков. Однако последний, вероятно, отличался отсутствием настоящей раковины, имел еще невысокое туловище, сходное с таковым у Lori-cata, и, по-видимому, вел более активный образ жизни, нежели Neopilina. От этого гипотетического первичного моллюска берут начало две главные линии развития. Одна ведет к Amphineura, другая — к Conchifera.
Среди боконервных наиболее примитивные черты: 8 спинных пластинок раковины, хорошо развитая радула, множественные жабры характерны для Loricata, тогда как Solenogastres, вероятно, представляют их вторично измененных потомков (утрата ноги и мантии, редукция раковины, радулы и т. д.).
У самого корня ветви Conchifera находится класс Monoplacophora, объединяющий некоторые существенные признаки Lamellibranchia, Gastropoda и Cephalopoda. Кристаллический стебелек кроме Neopilina имеется у многих пластинчатожаберных и низших брюхоногих. Общее
сходство развития сердца у Neopilina и высших Conchifera уже отмечалось. На филогенетическую связь с Cephalopoda (Tetrabranchia) указывают остатки метамерии у Nautilus. Нервная система в виде продольных тяжей свойственна низшим Gastropoda и Nautilus.
Таким образом, многое свидетельствует о происхождении высших классов Conchifera от форм, близких к Monoplacophorn. Однако современная Neopilina испытала уже некоторую специализацию, приспособившись к почти сидячему малоподвижному образу жизни.
От примитивных Monoplacophora, вероятно, берет свое начало гипотетическая группа билатерально симметричных моллюсков (так называемые Prorhipidoglossa), соединявшая в себе, по мнению многих зоологов, наиболее примитивные черты строения современных Gastropoda и Lamellibranchia. От этой группы расходятся в виде веера классы Gastropoda, Lamellibranchia и Scaphopoda, постепенно удаляющиеся друг от друга в лице своих высших и наиболее специализированных представителей. Обособление брюхоногих базируется на развитии у них асимметрии, а эволюция пластинчатожаберных характеризуется главным образом редукцией головы и образованием двустворчатой раковины.
Класс Cephalopoda стоит особняком, почему его приходится производить прямо от первичных моллюсков, близких к Monoplacophora, независимо от прочих классов Conchifera. Среди Cephalopoda подкласс че-тырехжаберных обладает, безусловно, более древними и примитивными чертами по сравнению с подклассом двужаберных.
Существует, однако, и другая точка зрения на происхождение и филогению моллюсков. Ее сторонники считают, что моллюски берут начало непосредственно от плоских червей (кл. Turbellaria), а самыми примитивными формами являются бороздчатобрюхие (кл. Solenogastres), объединяемые в отр. Chaetodermatina (или Caudofoveata). Простота строения в этом случае оказывается первичной, а не возникает в результате вторичного упрощения. Соответственно моллюски должны рассматриваться как исходно несегментированные животные. Их целом, представленный главным образом перикардием и отделяющимися от него в процессе индивидуального развития гонадами, не гомологичен вторичной полости тела кольчатых червей и возникает в процессе эволюции независимо от последней. Увеличение же числа отдельных органов и упорядоченность их расположения — вторичное явление, которое можно рассматривать как псевдометамеризацию.
Окончательное решение вопроса о природе моллюсков и их положении в системе животного царства, несомненно, требуют новых углубленных исследований.
ТИП ЩУПАЛЬЦЕВЫЕ (TENTACULATA)
Щупальцевые составляют группу вторичнополостных, олигомерных (малосегментных) животных не вполне выясненного происхождения. Щупальцевые ведут сидячий образ жизни, который оказал глубокое влияние на их организацию. Рассмотрим характерные признаки типа.
1. Тело неясно расчленено и слагается из трех сегментов. Первый — предротовая лопасть, или эпистом. Второй сегмент несет ротовое отверстие. Он окружен ресничными щупальцами, которые служат для подгона пищевых частиц ко рту и для дыхания. Щупальца сидят на щупаль-ценосце, или лофофоре, в основе имеющем форму подковы, или расположены по кругу. Третий сегмент — собственно туловищный. В ряде случаев эпистомальный сегмент вторично редуцируется.
2. Полость тела представлена целомом, который в соответствии с сегментацией разделяется на три отдела: передний эпистомальный, средний— сравнительно узкий, посылающий каналы в лофофор и щупальца, и задний, самый обширный — туловищный целом. Обособленность отделов целома может быть не всегда хорошо выражена.
3. Анальное отверстие лежит недалеко от рта, кишечник образует петлю.
4. Имеется кровеносная система, которая может вторично редуци-ваться.
5. Органы выделения Днефридии) представлены одной или двумя парами ресничных воронрк, открывающихся в целом. Являются ли эти образования метанефридиями или целомодуктами, пока окончательно не установлено. Иногда органы выделения редуцированы.
6. Половые железы образуются в туловищном целоме.
7. Дробление яйца полное, не спирального типа, из яйца развивается планктонная личинка, более или менее напоминающая трохофору.
К типу Tentaculata относятся 3 класса: мшанки (Bryozoa), плечено-гие (Brachiopoda), форониды (Phoronidea). Кроме того, в качестве дополнения к типу мы помещаем класс внутрипорошицевых (Entoprocta), систематическое положение которых неопределенно.
КЛАСС I. МШАНКИ (BRYOZOA)
Класс мшанок содержит водных, преимущественно морских, реже пресноводных животных, ведущих сидячий образ жизни и образующих в результате бесполого размножения колонии. Чаще всего колонии бывают древовидными, ветвистыми. В некоторых случаях (Flustra) ветви колонии сплющиваются в одной плоскости и принимают характер пластинок, а сама колония становится похожей на пучок листьев. Если же колонии стелются по субстрату, то образуют на дне или на стеблях водорослей тонкие корочки. Каждая колония объединяет большое количество особей. Таким образом, по внешнему виду колонии мшанок больше всего напоминают гидроидных полипов. Размеры мшанок невелики: колонии измеряются несколькими сантиметрами, отдельные особи большей частью длиной не более 1 мм. Современных видов мшанок около 4000, ископаемых еще больше.
Для понимания природы мшанок следует рассмотреть строение отдельной особи.
Строение и физиология.Отдельные особи колонии мшанок (рис. 496) отчетливо подразделяются на два отдела, которые, однако, не являются сегментами. Передний конец особи свободно торчит в окружающую воду и несет на себе рот с венчиком щупалец вокруг. Задняя половина животного одета утолщенной кутикулой, производимой наружным эпителием тела, и имеет вид глубокой чашечки или мешочка. Эту часть животного нередко обозначают особым термином — цистид. Передняя половина мшанки лишена плотной кутикулы, нежная и при раздражении втягивается внутрь цистида; ее называют полипидом (рис. 497). Втягивание совершается при помощи двух особых мускулов-ретракторов. Они находятся по бокам кишечника и передним концом прикрепляются изнутри к стенке тела полипида по сторонам рта, а задним концом — к основанию цистида. При втягивании передней половины тела в цистид отдельные щупальца не вворачиваются, а весь венчик их уходит в глубь образующегося впячивання. Щупальца имеют вид длинных нежных выростов, в которые заходят каналы целома, и покрыты мерцательным эпителием;
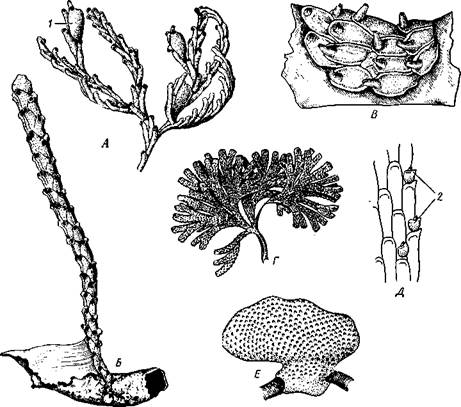
Рис. 495. Колонии морских мшанок подкласса Gymnolaemata. А — Idmonea tu-mida; Б — Crisia eburnea; В — Alcyonidium mamillatum; Г — Dendrobaenia flustrot-des; Д — участок колонии Dendrobaenia flustroides; E — Porella saccata: 1 — ГОНОЗОИД. 2 — авикулярия
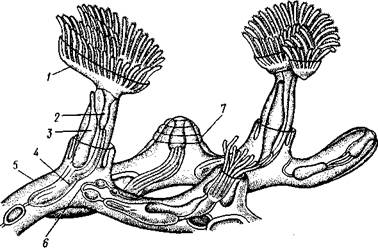
Рис. 496. Участок колонии Plumatella repens (из Матвеева)?
/ — полипид с расправленным лофофором, 1 — передний отдел кишечника, 3 — задняя кишка, 4 — желудок, 5 — стенка цистида, 6 — канатик со статобластамн.
7 — втянутый полипид
они служат, с одной стороны, для собирания пищевых частиц, с другой — в качестве органов дыхания.
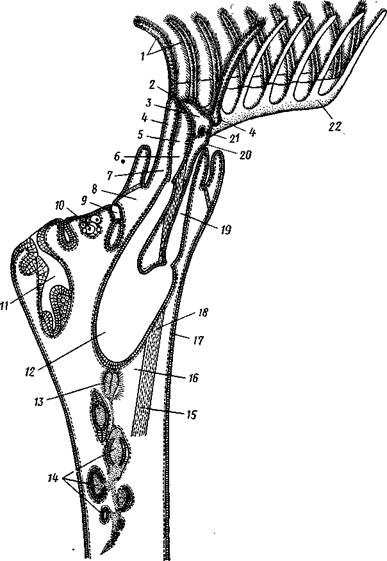
У пресноводных мшанок, образующих подкласс Phylactolaemata (Покрыторотые), щупальца в два ряда располагаются на особом щу-пальценосце — лофофоре, имеющем сверху, со стороны рта, форму под-ко.вы, у вершины которой и располагается ротовое отверстие (рис. 496, 497). Последнее прикрыто небольшим язычковидным выростом — эпи-стомом — предротовым отделом тела (рис. 497).
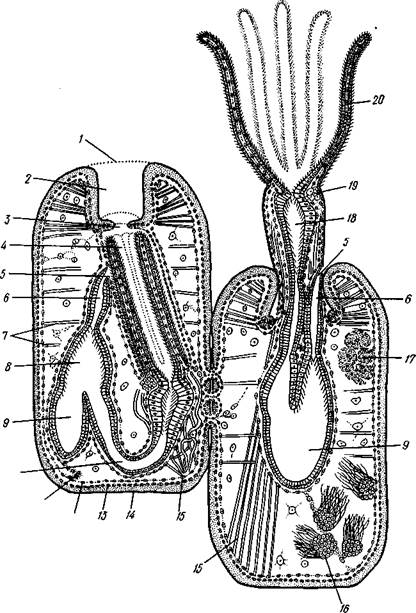
Представители подкласса Gymnolaemata (см. рис. 495) (Голоротые мшанки) вторично утратили эпистом, а их щупальце вокруг ротового отверстия образуют простой венчик (рис. 498).
Кутикула цистида может быть различной консистенции: в виде тонкой хитиновой пленки или остуденяется и превращается в толстый желеобразный слой, содержащий до 99% воды. В других случаях кутикула
|
| Рис. 497. Строение особи пресноводной мшанки из подкл. Phylactolaemata (из Ремане): I — щупальца, 2 — эпистом. 3 —* целом эпистома, 4 — целом ло-фофора, S—глотка, 6 — пищевод, 7 — полипид, 8 — цистид, 9 — оэций, 10 — яичник, 11 — почка, 12 — желудок, 13 — семенник, 14 — статобласт, 15 — канатик, 16 — туловищный целом, 17 — целомический эпителий, 18 — ретрактор, 19 — задняя кишка, 20 — анальное отверстие, 21 — ганглий, 22 — ло* фофор |
|
| ID |
| 11 |
| 12 |
| I |
Рис. 498. Схематическое строение двух зооидов мшанок отряда Gymno-laemata (в расправленном и втянутом состоянии) (из Маркуса):
/ — отверстие цистида, 2 и 3 — части устьевой области цистида, 4 — венчик щупалец, втянутый внутрь цистида, 5 — анус, б — задняя кишка, 7 — мускульные волокна, пересекающие целом, 8, 9 — желудок, 10 — пищевод, // — канатик, 12 — перитонеаль-ный эпителий, 13 — стенка тела, 14 — кутикула, 15 — мускул-ретрактор, 16 — семенники, 17 — яичник, 18 — глотка, 19 — нервный узел, 20 — щупальца
утолщается и пропитывается углекислой известью, становясь очень твердой.
Наружный эпителий мшанок однослоен и у пресноводных форм (подкл. Phylactolaemata) подстилается двумя слабо развитыми слоями мышц: наружным из кольцевых и внутренним из продольных мышечных волокон. У морских голоротых мшанок (подкл. Gymnolaemata) мышечные слои отсутствуют. Слабое развитие кожно-мускульного мешка объясняется неподвижностью мшанок. Перитонеальный эпителий ограничивает обширную вторичную полость тела, в которой помещаются все внутренности.
Полость тела (целом) тонкими перегородками делится на три отдела. Передний обладает небольшими размерами и залегает в эпистоме. Средний — кольцевой канал — окружает глотку и посылает слепые ответвления в щупальца. Задний, наиболее обширный, занимает почти все тело и называется туловищным целомом. Как уже говорилось, у голоротых мшанок эпистом вместе с целомом редуцируется.
Пищеварительная система. Пищеварительный канал имеет характерную для многих сидячих животных подковообразную форму (см. рис. 496, 497, 498). Рот, лежащий внутри венчика щупалец (у подкл. Phylactotaemata— в средней части подковы лофофора, между двумя рядами ее щупалец), ведет сначала в небольшую глотку, а затем в длинную узкую трубку — пищевод. Пищевод переходит в мешковидный, V-образно изогнутый желудок; он впадает в одно из колен желудка, тогда как от другого колена отходит поднимающаяся кпереди тонкая кишка, открывающаяся наружу порошицей непосредственно позади венчика щупалец.
Весь кишечник снаружи одет перитонеальным эпителием. От дна желудка отходит продолжение его перитонеальной выстилки в виде тонкой брыжейки, или канатика, тянущегося через полость тела назад и переходящего затем в наружный, пристенный слой перитонеума.
Пища — мелкие одноклеточные водоросли, простейшие, коловратки и т. п. — подгоняется ко рту мерцанием ресниц на щупальцах. В глотке пища сбивается в пищевые комки и поступает через пищевод в желудок.
Нервная система состоит из единственного надглоточного ганглия (см. рис. 497, 498), лежащего между глоткой и задней кишкой, от которого расходятся в разные стороны, но главным образом к щупальцам, периферические нервы. Единственными органами чувств являются особые осязательные волоски на наружной стороне щупалец.
Интересно, что во время почкования ганглий почки возникает в виде ясного впячивания эктодермы, отшнуровывающегося от покровов; полость впячивания позднее исчезает, и ганглий становится массивным.
Дыхание происходит через всю поверхность тела, но главным образом через щупальца.
Кровеносной системы у мшанок нет. Отсутствие ее, вероятно, объясняется редукцией, вызванной малыми размерами тела, что, в свою очередь, связано с тем, что мшанки — животные колониальные.
Выделительные органы у мшанок отсутствуют. Удаление продуктов обмена из организма осуществляется с помощью клеток-фагоцитов через стенки щупалец или стенку кишечника. Из последнего они выводятся вместе с экскрементами.
У покрыторотых мшанок полости целомов соединяются трубчатыми образованиями, которые рассматриваются как видоизмененные целомо-дукты. Долгое время их считали органами выделения покрыторотых мшанок, но сейчас эта точка зрения оставлена.
Половая система. Мшанки — гермафродиты. Половые клетки раз-
виваются под перитонеальным эпителием, на стенке тела или на канатике. Живчики образуют на канатике довольно большое скопление — семенник. Яйца формируются на внутренней стенке тела (рис. 498) в небольшом количестве, иногда по одному. Оплодотворение внутреннее — подвижные живчики выходят из одной колонии мшанок и активно проникают в другую, где в это время имеются созревшие яйца. Выход оплодотворенных яиц наружу совершается разными способами. У одних видов для этого используется специальная целомическая пора, расположенная между щупальцами и ротовым отверстием. Значительно чаще развивается специальный ресничный канал, который открывается наружу непосредственно между щупальцами. Наконец, как полагают, у ряда видов зрелые яйца обладают активным амебоидным движением и сами прокладывают себе дорогу через стенки родительской особи.
У очень немногих видов оплодотворенные яйца выбрасываются в воду, где и осуществляется их дальнейшее развитие. Более обычно проявление у мшанок своеобразной «заботы о потомстве», когда яйца приклеиваются к покровам или даже поступают в особые образования — оэции (см. ниже), в которых протекает их развитие до полного формирования личинки.
Колонии. У мшанок, как и у многих других колониальных организмов, наблюдается морфологическая и функциональная дифференци-ровка особей, образующих колонию. У сохранивших большое количество примитивных признаков покрыторотых мшанок в колониях представлены особи только двух типов. Основную массу составляют обычные индивиды. Кроме них имеются еще оэции, которые образуются за счет молодых почек, развитие которых рано останавливается. Почка у Phy-lactolaemata сначала представляет небольшое впячивание покровов, в полость которого и поступает яйцо. Оэции с развивающимся зародышем до конца сохраняет вид простого мешочка. В колониях подкл. Gymnolae-mata дифференциация особей выражена значительно сильнее. Оэции го-лоротых мшанок имеют вид выступающих бугорков, несущих полость, сообщающуюся с внешней средой. В этой полости протекает формирование личинки. У некоторых видов отдельные особи колонии резко увеличиваются в размерах и принимают характерную кувшинообразную форму. Они называются гонозоидами (см. рис. 495). В полости гонозоидов яйца претерпевают сложное развитие, включающее полиэмбрионию (с. 509).
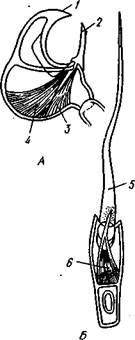
Ряд особей колонии специализируется для выполнения защитных функций. Это в первую очередь авикулярии (рис. 499). Последние имеют вид, сходный с птичьей головой (с шеей и мощным клювом). «Голова» представляет собой цистид, измененный и вытянутый в неподвижный отросток, отвечающий верхней части «клюва». Кроме того, на «голове» сидит подвижный кутикулярный крючок, отвечающий нижней части клюва; он способен то прижиматься к неподвижному отростку, то отводиться от него. Подвижный кутикулярный крючок есть результат видоизменения крышечки, имеющейся у многих мшанок на переднем крае цистида и захлопывающейся при втягивании полипида. Авикулярии захватывают мелких животных, заползающих на колонию, и убивают или прогоняют их. Разновидностью авикулярии являются вибракулы, которые отличаются отсутствием неподвижного выроста цистида, тогда как кутикулярный крючок вытягивается в длинный подвижный жгут.
Размножение.Мшанки имеют половое размножение, но, кроме того, отличаются большим разнообразием бесполых способов размножения. Прежде всего образование колоний — результат не доведенного до конца бесполого размножения посредством почкования. Детали процесса
|
почкования у мшанок довольно сложны, но в общем он состоит в том, что на поверхности колонии образуются в определенных (у разных родов кл. Bryozoa различных) местах бугорки — зачатки будущих почек. Бугорок растет, и перетягивается у основания, но полностью не отделяется; почка остается в связи с колонией. На почке путем впячивания отдельных участков ее эктодермы закладываются кишечник и ганглий. Щупальца закладываются тоже в виде впячиваний, которые потом выворачиваются наружу. Соответственно месту образования почек на материнском организме, количеству их и дальнейшему способу почкования колония мшанок принимает различную форму. Почки формируются во вполне развитые особи и, в свою очередь, начинают размножаться почкованием.
В результате такого наружного почкования происходит постепенный рост колонии.

Рис. 499. Авикулярия (А) и вибракула (Б) мшанки (из
Догеля):
/ — неподвижная ветвь, 2 — подвижная ветвь, 3, 4—мышцы, раскрывающие и закрывающие «клюв», 5 — подвижный бич вибракулы, приводимый в действие мышцами (6)
Рис. 500. Статобласт Crista-' tella mucedo. Вокруг цент-' ральной массы видно плава-• тельное кольцо, состоящее > из хитиновых камер, наполненных газом, и хитиноид-ные крючочки (по Догелю)
Наряду с наружным почкованием у пресноводных мшанок (отр. Phylactolaemata) имеется еще особый способ внутреннего почкования при помощи образования статобластов (см. рис. 497; 500). Статоблас-ты — особые чечевицеобразные многоклеточные тельца, одетые плотной оболочкой; образуются преимущественно осенью. Зачаток статобласта появляется внутри канатика в виде кучки мезодермальных клеток. Одновременно с этим группа эктодермальных клеток мигрирует с поверхности тела внутрь канатика. Здесь за их счет формируется двухслойная эпителиальная оболочка, окружающая группу мезодермальных клеток. Эпителий выделяет на своей поверхности тонкую, но плотную скорлупу, состоящую из двух слоев, между которыми располагается слой воздухоносных камер. У некоторых видов по краю чечевицы образуется хитиновое кольцо с крючками, или зацепками. Статобласты освобождаются при разрушении материнского тела (например, зимой при отмирании колонии) и попадают наружу. Воздухоносные камеры позволяют им плавать в воде, а своими крючками они зацепляются за водоросли, неровности дна и т. п. Статобласты остаются зимой в покоящемся состоянии,
весной же оболочки их лопаются, клеточное содержимое прикрепляется ко дну и образует при помощи наружного почкования новую колонию.
Легко видеть, что внутреннее почкование мшанок имеет большое сходство и одинаковое биологическое значение с образованием геммул у пресноводных губок (с. 108), Наконец, у некоторых мшанок, относящихся к подкл. Gymnolaemata, зимуют особые «зимние» наружные почки: они представляют собой одетые плотной оболочкой отдельные цистиды. От статобластов они легко отличаются по присутствию в них зачатка кишечника, мышц и половых клеток, тогда как внутренность статобласта занята сплошной массой мезодермальных клеток.
Развитие.Все мшанки обладают половым размножением. Дробление яйца полное и во многих случаях почти равномерное. В результате возникает бластула, часто сплющенная в одном направлении. В этом случае она сначала имеет вид пластинки, а позднее становится похожей на чечевицу.
Часть клеток на одном полюсе бластулы внедряется в бластоцель и дает начало неразделенному зачатку энтодермы и мезодермы.
У многих видов, развитие эмбрионов которых протекает в оэции или гонозоиде, между формирующимся зародышем и материнской колонией устанавливается тесная связь в виде своеобразной «плаценты». Таким способом эмбрион получает необходимые для развития питательные вещества.
Процессы эмбрионального развития части мшанок из подкл. Gymnolaemata усложняются за счет появления полиэмбрионии. Оплодотворенное яйцо, развивающееся в гонозоиде, в результате неравномерного дробления дает сначала крупного первичного зародыша, от которого позднее отделяются более мелкие вторичные зародыши. Количество последних может достигать ста и более. В результате развития получаются свободноплавающие личинки неодинакового вида у разных мшанок (рис. 501). Наиболее типичная личинка, сохранившая ряд примитивных признаков,— цифонаут; тело ее заключено в двустворчатую раковину. Передвигается личинка с помощью венчика ресничек. Имеется хорошо развитый кишечник; на переднем конце перед ротовым отверстием располагается особый грушевидный орган, выполняющий функции органа чувств, а на брюшной поверхности — присоска, с помощью которой личинка прикрепляется к субстрату во время метаморфоза.
У многих мшанок наблюдается вторичное упрощение личинок, связанное с утратой раковины и кишечника. Самостоятельный энтодермальный зачаток в этом случае не обособляется.
После периода свободного плавания личинка садится на дно, прикрепляется и испытывает превращение. Тело осевшей личинки принимает вид овального мешочка, и большинство личиночных органов подвергается распаду, которому содействуют амебоидные клетки фагоцитарного характера. В частности, у цифонаута полностью разрушается энтодермальный кишечник. Новый кишечник формируется за счет экто-дермальных клеток. На свободной стороне личинки, противолежащей месту прикрепления, начинается закладка одной или двух первых особей будущей колонии. Они образуются в виде утолщений эктодермы, которые впячиваются внутрь и дают особь мшанки по способу, напоминающему наружное почкование. Путем дальнейшего почкования первых особей и их потомков постепенно вырастает целая колония.
У пресноводных мшанок (подкл. Phylactolaemata) закладка отдельных особей происходит очень рано — еще во время развития личинки. У Cristatella личинки, выходящие из оэция в воду, несут до 20 особей, находящихся на разных стадиях формирования.
Экология.Мшанки живут преимущественно в море, где встречаются на различных глубинах, начиная с прибрежных камней и до больших глубин.
В пресных водах встречаются почти исключительно снабженные щупальценосцем Phylactolaemata. Наиболее обыкновенны из них виды рода Plumatella, образующие стелющиеся по субстрату, ветвящиеся колонии или массивные наросты на подводных предметах, и Cristatella mucedo (рис. 502). Последняя особенно интересна тем, что у нее проявляется высокая степень интеграции колонии, выражающаяся в слиянии цистидов. Колонии Cristatella имеют вид студенистой колбаски, на верхней поверхности которой торчат полипиды. Подошва не прикреплена к субстрату, и вся колония может очень медленно ползать.
Срок жизни пресноводных колоний обычно не больше 5—6 месяцев; колонии редко перезимовывают. Жизнь отдельных особей колоний значительно короче; на колонии всегда можно встретить дегенерирующие особи, места которых впоследствии занимаются индивидами, вновь развивающимися из почек.
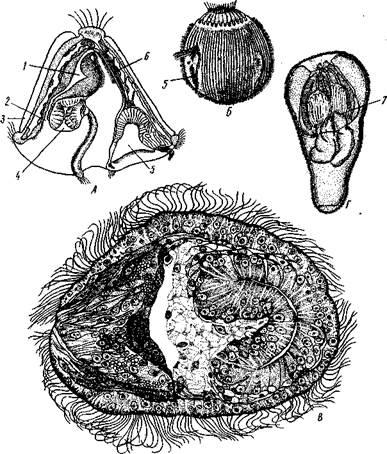
Рис. 501. Личинки мшанок (из разных авторов). А—личинка типа цифонаут; Б — личинка Bugula sp.; В — личинка Crisia sp.; Г — личинка Plumatella до выворачивания полипидов:
Дата добавления: 2015-01-24; просмотров: 2842;