Филогения типа Arthropoda
Происхождение типа Arthropoda в общих чертах ясно. Предками их были примитивные полимерные кольчатые черви из класса многощетин-ковых (Polychaeta). Сходство плана строения членистоногих и кольчецов настолько велико, что некоторые современные зоологи предпочитают объединять их, как это делали свыше 100 лет назад, в один тип членистых — Articulata.
По расчленению на отделы тела (тагмы), характеру и степени дифференциации головных и туловищных конечностей и по особенностям личиночных стадий подтипы членистоногих заметно отличаются друг от друга.
Переход от кольчецов к членистоногим сопровождался усложнением строения: превращением довольно тонкой кутикулы в жесткий наружный скелет, распадом кожно-мускульного мешка на отдельные мышцы, появлением смешанной полости тела (миксоцеля), превращением параподийв членистые конечности, спинного сосуда — в более резко обособленное сердце, развитием сложных глаз, а также постепенной заменой гомоном-пой сегментации тела гетерономной в процессе цефализацйи, т. е. превращения передних туловищных сегментов в головные, а их конечностей — в ротовые органы.
Часть членистоногих сохраняет первоначальный водный образ жизни кольчецов, сохраняет и органы водного дыхания, т. е. наружные кожные жабры. В связи с тем что жабры помещались у полихет на параподиях, они сохраняют то же положение и у низших представителей типа Arthro-
1 ч
poda с той разницей, что параподии расчленяются и превращаются в в настоящие ноги. Пальпы предковых форм, по-видимому, дают начало антеннулам членистоногих.
Наиболее примитивные формы членистоногих мы находим среди представителей подтипов Branchiata и Trilobitomorpha. Обе эти группы обладают рядом исходных первичных признаков, и тем не менее они резко отличаются друг от друга. В первую очередь эти различия касаются строения конечностей: у ракообразных они сохранили древнюю двувет-вистость, а у трилобитов претерпели специализацию и стали одновет-вистыми. С другой стороны, у трилобитов отсутствует морфологическая дифференциация ножек (и на туловищных и на головных сегментах они имеют одинаковое строение). В то же время даже у многих примитивных раков морфологическая и функциональная специализация конечностей выражена достаточно отчетливо. Далеко зашел у трилобитов и процесс цефализации — все они обладают вполне обособленной цельной головой, тогда как у низших представителей подтипа Branchiata сохраняется расчленение головного отдела на протоцефалон и свободные челюстные сегменты. Все это, несомненно, говорит о том, что трилобиты и ракообразные — две самостоятельные филогенетические ветви, очень рано обособившиеся от каких-то общих предков.
Branchiata — древняя группа членистоногих. Уже в кембрийских отложениях встречаются представители современных отрядов. У ракообразных четыре передних сегмента становятся головными, а их конечности соответственно дают антенны и три пары челюстей. Тем не менее цефализация у многих ракообразных остается еще на сравнительно низкой ступени — у отрядов Anostraca и Mystacocarida, а также у многих представителей подкл. Malacostraca имеется первичная голова — протоцефалоп.
Подтип Tracheata составляет еще одну группу членистоногих, тесно связанную с подтипом Branchiata, с которыми они, несмотря па наземный образ жизни, имеют очень много общего. В самом деле, в обеих группах дифференциация головных конечностей в сущности одинакова. Трахейные, правда, не имеют усиков II — антенн, но они утратили их вторично. В составе головы сохранился еще сегмент антенн с принадлежащими ему ганглиями, т. с. тритоцеребрумом. У зародышей некоторых трахейных были найдены зачатки антенн, которые позднее исчезают. Ротовые органы жабродышащих и трахейных сходны, в обоих случаях это верхние челюсти (мандибулы) и две нары нижних челюстей (мак-силл). Все это — свидетельства общности происхождения подтипов Branchiata и Tracheata и в какой-то мере оправдывает объединение их в большую группу жвалоносных, или челюстных, членистоногих (Мап-dibulata).
Вторая филогенетическая ветвь членистоногих берет начало от Trilobitomorpha. Последние просуществовали до конца палеозоя, оставив видоизмененных потомков — хелицеровых (Chelicerata); последние утратили усики-антеннулы; первая пара их головных ножек дифференцировалась в клешненосные хелицеры, вторая — в педипальпы, остальные две пары превратились в две передние пары ходных ног. Как и ходные ноги, дифференцировались также конечности двух передних сегментов туловища, которые у большинства хелицеровых слились с головой, образовав головогрудь. Остальные сегменты образовали передне- и заднебрю-шие, причем на первом конечности долгое время сохраняли дыхательную функцию, тогда как на втором — редуцировались. Таким образом, Chelicerata связаны с Polychaeta через трилобитов; низшие классы хелицеровых, подобно своим предкам, — водные животные с жаберным дыха-
нием, высший класс — паукообразные (кл. Arachnida)—приспособился к жизни на суше. Особенности специализации конечностей головных сегментов у трилобитообразных и хелицеровых позволяют их также объединять в большую группу Amandibulata — бесчелюстных членистоногих.
Таким образом, среди членистоногих следует различать по крайней мере две большие очень рано обособившиеся филогенетические ветви: Mandibulata (Branchiata — Tracheata) и Amandibulata (Trilobitomor-pha,— Chelicerata).
1 Так как аннелидные предки типа Arthropoda обладали неопределенно большим числом гомономных сегментов, то естественно, что эта черта строения характерна и для наиболее примитивных членистоногих. Прослеживая эволюцию отдельных ветвей типа Arthropoda, мы видим ряд параллельно протекающих процессов. Одни из них суть характерные явления, сопровождающие всякую прогрессивную эволюцию: это дифференциация частей и олигомеризация органов, а также интеграция отделов тела и всего организма. Так, во всех ветвях членистоногих мы неизменно видим, как постепенно число сегментов уменьшается, т. е. происходит их олигомеризация. Параллельно с этим гетерономность сегментации возрастает, происходит дифференциация самих сегментов, конечностей и вначале метамерных внутренних органов. Другие изменения объясняются общностью происхождения — в ветвях, дивергирующих друг от друга, тем не менее переход к одинаковому образу жизни (например, наземному) приводит к сходным изменениям (появление трахей, развитие мальпигиевых сосудов и др.).
Перейдем к рассмотрению филогенетических отношений внутри подтипов членистоногих.
Branchiata.Среди ракообразных примитивные признаки сохраняются полнее всего у отряда жаброногих (Anostraca): туловище их гомономно расчленено. Антенны иннервируются от окологлоточных коннективов, нервная система имеет характер брюшной лестницы и мало концентрирована, сердце длинное, трубчатое. Короткие конечности двуветвисты, сохраняется первичная голова — протоцефалон. Наконец, число сегментов у жаброногих очень велико — до 31 (Branchipus и др.). Велико оно и в другом очень примитивном отряде листоногих раков (Phyllopoda), в частности в подотряде щитней (Notostraca) достигает 46; у раковинных листоногих раков (подотряд Conchostraca) в состав тела входит 36—14 сегментов; среди этих ракообразных есть формы, во многих отношениях приближающиеся к подотряду ветвистоусых (Cladocera), примитивные представители которых (Sida) имеют только 10 сегментов (считая и входящие в состав головы), у прочих же ветвистоусых число сегментов падает до 9, а у специализированного Polyphaemus — даже до 8.
Примитивны и цефалокариды, сохранившие ряд признаков, характерных, по-видимому, для предков всех ракообразных.
Другие линии эволюции ракообразных представляют подклассы ра-кушковых (Ostracoda) и максиллопод (Maxillopoda). Эти специализированные в разных направлениях группы, несомненно, тоже произошедшие от многосегментных предков (хотя точные филогенетические связи их неясны), обладают небольшим числом сегментов (Ostracoda — менее 10, Maxillopoda—14). Таким образом, у ракообразных мы видим в разных филогенетических ветвях независимую и параллельную олиго-меризацию сегментов.
Несомненно, подкл. Malacostraca не находится в непосредственном родстве с другими подклассами, так как в некоторых отношениях они примитивнее остальных ракообразных (наличие у них брюшных ножек,
сохранение первичной головы — протоцефалона, свойственной лишь самым низшим группа^ раков, и т. п.). Поэтому надо считать, что подкласс Malacostraca берет начало независимо от других подклассов от общих для всех ракообразных примитивных и отдаленных предков. Среди высших раков наиболее примитивен отряд тонкопанцирных (Lepto-straca). Об этом можно судить по присутствию у взрослых раков двух пар целомодуктов — антеннальных и максиллярных желез, наличию седьмого сегмента брюшка и т. д., напротив, отряды Isopoda и Amphi-poda отличаются далеко зашедшей цефализацией, у них сложная голова, к которой присоединились 1—2 сегмента груди.
Tracheata.Вопрос о филогенетических взаимоотношениях внутри класса Myriapoda в настоящее время не может считаться окончательно решенным. Одни исследователи рассматривают отдельные группы многоножек как самостоятельные, рано обособившиеся классы членистоногих. Другие считают, что для этого нет достаточных оснований и более правильно всех многоножек объединить. Наиболее примитивными из Myriapoda, по'-видимому, нужно считать губоногих (Chilopoda), о чем свидетельствует ряд признаков, и, в первую очередь, большое и непостоянное в пределах группы число сегментов (до 177) и гомономность, отчетливо выраженная у ряда низших форм. Вторичные изменения сильнее всего проявляются у двупарноногих, или кивсяков (Diplopoda), у которых произошло попарное слияние большинства туловищных сегментов.
"" На происхождение насекомых имеются два взгляда. Одни ученые, точки зрения которых придерживаемся и мы, ведут генеалогию насекомых от многоножек. Другие, к которым принадлежат в особенности палеонтологи, производят насекомых непосредственно от трилобитов или от древнейших раков. Остановимся на второй точке зрения. Она основана прежде всего на том, что почти все самые древние из известных нам ископаемых насекомых, а именно каменноугольные, относятся к группам, похожим на современных поденок, веснянок и стрекоз, которые ведут земноводный образ жизни, но личинки их живут в воде. Отсюда делается вывод, что они представляют собой как бы переход от древних водных членистоногих к предкам современных насекомых. Трахейные жабры личинок упомянутых насекомых рассматриваются как видоизмененные конечности трилобитных предков. Не умаляя значения палеонтологической летописи, мы, однако, полагаем, что, судя по раннему обособлению почти всех других беспозвоночных, настоящих предков насекомых надо искать где-нибудь глубже каменноугольных слоев. Кроме того, все насекомые дышат трахеями; между тем трахеи возникают всегда в виде впячиваний покровов и притом у членистоногих, уже прочно перешедших к наземному образу жизни. Поэтому трахейные жабры поденок и веснянок могут расцениваться как вторично приобретенное приспособление к водному дыханию наземных животных, вторично перешедших к жизни в воде. Следовательно, и земноводный характер каменноугольных насекомых вторичен.
Поэтому мы предпочитаем держаться первого взгляда на филогению насекомых. Насекомые ведут начало от каких-то многоножкообразных предков. Ни одну из современных групп многоножек нельзя признать предками насекомых, но совокупность признаков, имеющихся у разных представителей многоножек, поддерживает высказанную точку зрения. Большое количество сегментов тела, более гомономная сегментация, присутствие ходных ножек почти на всех сегментах тела — все эти признаки доказывают большую примитивность многоножек по сравнению с насекомыми.
Взаимоотношения отдельных групп в пределах самого класса насекомых не столь ясны. У представителей подкласса Entognatha сохраняется ряд примитивных черт организации — богатое расчленение тела (брюшко Protura обладает исходным для насекомых числом сегментов— 11), наличие рудиментарных или сильно видоизмененных конечностей на части брюшных сегментов, первичное отсутствие крыльев, развитие с анаморфозом. Однако наряду с этим у них в процессе эволюции появились и признаки далеко зашедшей специализации — образование ротовой капсулы, в которую погружаются ротовые части, возникновение колющих и сосущих ротовых аппаратов и т. п. Мы не склонны рассматривать скрыточелюстных в качестве самостоятельного класса и тем более как ряд самостоятельных классов, хотя подобная точка зрения и существует в современной зоологической литературе. Однако необходимо признать, что подкласс Entognatha довольно рано отделился от общего ствола насекомых и прошел свой длительный и самостоятельный путь эволюции. Сохранению же целого комплекса примитивных признаков, встречающегося у представителей этой группы, способствовало обитание в почве и скрытный образ жизни.
Среди представителей подкласса Ectognatha, не утративших исходное первичное положение ротовых конечностей на поверхности головы, наиболее примитивными, несомненно, являются тизануры (отр. Thysa-nura), у которых некоторые исследователи обнаруживают ряд признаков, роднящих их с многоножками.
Что касается олигомеризации у Tracheata, то она ярче всего проявляется в числе сегментов, конечностей, трахей и дыхалец. Многоножки обладают большей частью многочисленными сегментами, количество которых сильно варьирует в разных группах. Но для высшего класса Tracheata насекомых характерно заметное уменьшение количества сегментов. Лишь у протур 18 сегментов (считая 4 головных). У других< форм эта цифра сокращается до 17. Тело многих высших насекомых' состоит всего из 11 —12 сегментов.
Trilobitomorpha — Chelicerata. Трилобиты — один из наиболее примитивных классов членистоногих. Самые древние трилобиты из кембрия отличаются расчлененным и гомономным туловищем, напротив, более поздние обнаруживают уменьшение числа туловищных сегментов и слияние задних сегментов в хвостовой щит. Максимального расцвета трилобиты достигли в нижнем силуре, число семейств дошло до 77 при большом количестве видов с очень разнообразной биологией. Начиная с верхнего силура идет постепенное вымирание трилобитов, и уже в пермский период они исчезают. Приблизительно в одно время с трилобитами развиваются и произошедшие от них древние водные формы подтипа Cheli-ccrata, а именно класса Gigantostraca, появляющиеся в кембрии и вымирающие в пермский период. Большой древностью отличается и другой класс хелицеровых — Xiphosura, дожившие с силура до наших дней. С того же времени становятся известны и паукообразные, а именно очень близкий к современным скорпионам, но живший в воде Palaeophonus. Однако сильного развития сухопутные хелицеровые достигли лишь в карбоне.
Оба низших класса подтипа Chelicerata в отличие от многосегментных трилобитных предков имеют уменьшенное и стабилизированное число сегментов: класс Gigantostraca обладает 19 сегментами, а у класса Xiphosura часть задних сегментов выпадает совсем, часть входит в состав хвостового шипа (с. 403), так что у взрослых животных туловище состоит лишь из 13 сегментов. Антеннулы редуцируются.
Класс паукообразных теснее всего связан с классом Gigantostraca.
У них одинаковое количество, а отчасти форма и функция конечностей головогруди, происхождение легких от жаброносных брюшных ножек и отсутствие у обеих групп антеннул и ряд других общих признаков. Силурийский скорпион — Palaeophonus настолько еще сходен с некоторыми Gigantostraca, что может рассматриваться как настоящая переходная форма.
Среди паукообразных наиболее примитивные отношения сохраняются у богато расчлененных скорпионов и жгутоногих, а также может быть у сольпуг и некоторых низших клещей. По мере эволюции отдельных ветвей Arachnida наблюдается олигомеризация числа сегментов. Скорпионы имеют в составе тела 19 сегментов, жгутоногие и ложные скорпионы— 18, примитивные членистобрюхие пауки—17, сольпуги и сенокосцы— 16, большинство клещей—13, высшие пауки 12 и, наконец, некоторые специализированные клещи—10. Общим признаком хелице-ровых является отчетливо выраженная у них тенденция к рудиментации или полному исчезновению VII сегмента (с. 401). Параллельно происходит слияние сегментов и концентрация отделов тела. Гомологи брюшных конечностей, свойственные некоторым отрядам, — половые крышечки, гребневидные органы, легкие, паутинные бородавки — также подвергаются олигомеризации. Так, например, примитивные скорпионы имеют 4 пары легких, низшие пауки и жгутоногие — 2 пары, высшие пауки — 1 пару, у остальных Arachnida легкие совсем исчезают и полностью замещаются трахеями.
Рассмотрим взаимоот'ношения трех сравнительно небольших групп: кл. Pantopoda, кл. Tardigrada и кл. Linguatulida и остальных представителей типа Arthropoda. Каждая из этих групп обладает рядом специфических особенностей, которые не позволяют безоговорочно относить их ни к одной из известных групп членистоногих.
Морские пауки (кл. Pantopoda), по-видимому, ближе других стоят к хелицеровым. С последними они сближаются отсутствием усиков (антенн), наличием хелифор и пальп, которые очень похожи на хелицеры и педипальпы, далее — развитием ног на грудном отделе и редукцией их на брюшке. Хелифоры иннервируются, так же как хелицеры, от трито-церебрального отдела мозга. Некоторое сходство с паукообразными проявляется в общем виде, в форме головных конечностей и в наличии паутинных желез. Делались попытки сближать морских пауков с ракообразными на основании некоторого сходства между личинками, прото-нимфоном и науплиусом (с. 426) и общих черт метаморфоза. Однако в остальном морские пауки явно очень далеки и от хелицеровых, и от ракообразных, что особенно ярко проявляется в особенностях сегментации этих животных (с. 293, 402, 425). Некоторые исследователи считают, что морские пауки — это самостоятельная эволюционная веточка, берущая начало от каких-то древних предковых форм членистоногих.
Еще более неясно систематическое положение тихоходок. Сделано много попыток сближения кл. Tardigrada с различными группами: коловратками, нематодами, низшими ракообразными и даже с регрессировавшими неотеническими формами личинок насекомых. Наличие 4 пар ходных ног и толкование стилетов как измененных хелицер заставляло некоторых зоологов считать тихоходок одним из отрядов паукообразных.
В настоящее время правильнее всего считать тихоходок за самостоятельный примитивный класс, близкий к членистоногим, который, возможно, независимо от членистоногих ведет происхождение от многоше-тинковых червей. Конечности тихоходок еще не перешагнули через ступень сложности строения, свойственную параподиям полихет, коготки вполне похожи на некоторые формы щетинок; нервная система своею
гомономностью даже ближе стоит к аннелидам, чем к членистоногим. С другой стороны, полная кутикуляризация покровов и стенок передней и задней кишок, отсутствие ресничек, наличие линек, малышгиевы сосуды, если за таковые можно принять выросты задней части кишки, — признаки, свойственные членистоногим.
Столь же затруднительно точное определение положения в системе кл. Linguatulida. Подавляющее большинство признаков как будто говорит в пользу отнесения их к типу Arthropoda, а именно к подтипу Che-licerata, но окончательное решение вопроса затруднено отсутствием выделительной, кровеносной и дыхательной систем. О принадлежности к членистоногим свидетельствует гистологическое строение: хитиновая кутикула, поперечнополосатые мышцы, отсутствие ресничек. Сближение с хелицеровыми членистоногими обосновывается некоторым сходством с частью клещей по форме тела и конечностей. Кроме того, некоторые клещи паразитируют в дыхательных органах позвоночных, и как раз в отряде клещей мы видим тенденцию к утрате дыхательной и кровеносной систем.
Однако наличие нерасчлененных простых конечностей не позволяет относить кл. Linguatulida к настоящим членистоногим и сближает их с кл. Tardigrada. Все это заставляет считать перечисленные группы как дополнения к типу Arthropoda.
ТИП ОНИХОФОРЫ (ONYCHOPHORA)
Онихофоры — небольшая группа (70 видов) червеобразных, сегментированных наземных животных, обитающих в тропиках и умеренных странах южного полушария. Прежде их причисляли к членистоногим, именно к подтипу Tracheata, под'названием Первичнотрахейных (Рго-tracheata) и рассматривали как промежуточное звено между кольчатыми червями и многоножками. Строение и эмбриональное развитие они-хофор показывают, что общие с подтипом Tracheata черты выработались у них независимо в результате параллельного эволюционного развития. Поэтому в настоящее время Onychophora рассматриваются как самостоятельный тип сегментированных первичноротых животных.
Для онихофор характерны следующие особенности.
1. Тело неясно разделяется на голову с усиками и длинное очень гомономное туловище без границ между сегментами, но с парными конечностями примитивного строения.
2. Наружный хитиновый скелет отсутствует, имеется кожно-мускуль-ный мешок, состоящий из гладкой мускулатуры.
3. Полость тела смешанная (миксоцель).
4. Незамкнутая кровеносная система представлена лежащим на спинной стороне тела сердцем с метамерными остиями.
5. Органы выделения представлены почти во всех сегментах мета-мерными целомодуктами.
6. Дыхание осуществляется посредством воздухоносных трубочек — трахей.
К типу онихофор принадлежит один класс — Первичнотрахейные (Protracheata).
КЛАСС I. ПЕРВИЧНОТРАХЕЙНЫЕ (PROTRACHEATA)
Все первичнртрахейные внешне напоминают кольчатых червей или крупных медлительных голых гусениц. Большое наружное сходство Protracheata с некоторыми формами Polychaeta (например, Hesione)
одно время заставляло считать их особым семейством многощетинковых кольчатых червей.
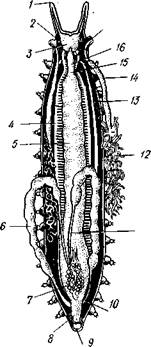
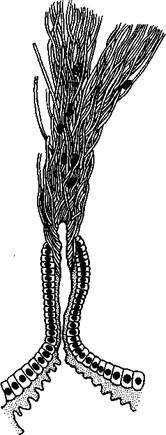
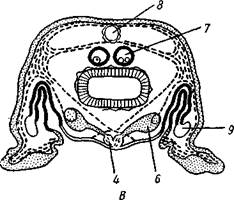
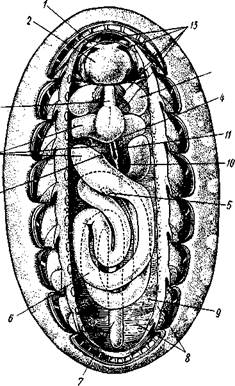
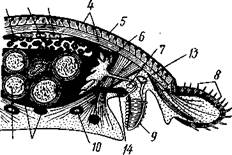
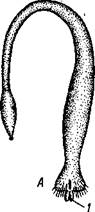
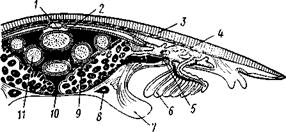
Строение и физиология.Длинное (до 15 см) цилиндрическое тело (рис. 418) состоит из ряда (от 13 до 43) сегментов, снабженных каждый парой ножек. Голова (рис. 419) состоит из трех сегментов, присоединившихся к головной лопасти (акрону); на спинной ее стороне, впереди рта, выдаются длинные антенны, у основания которых лежит пара глаз.

Рис. 418. Перипатопсис Peripatopsis capensis (по Седжвику)
Антенны иннервируются от головного мозга и отвечают первой паре усиков (антеннулам) раков и единственной паре антенн многоножек и насекомых (с. 333, 341). Антенны имеют поверхностную кольчатость, но в действительности не расчленены. Глаза и антенны принадлежат головной лопасти — акрону. Первый сегмент головы, вставочный или интерка-лярный, лишен придатков, но имеет соответствующий ему отдел головного мозга. Второй головной сегмент несет конечности, превратившиеся в ротовые органы. В ротовой полости лежат два мускулистых бугорка, снабженных каждый хитиновой зазубренной пластинкой. Это челюсти, гомологичные ходным ножкам, которые переместились в ротовую полость и приняли на себя функцию размельчения пищи. Третий сегмент головы также имеет пару измененных ножек в виде ротовых сосочков, расположенных по бокам головы и несущих на своих вершинах отверстие особых слизистых кожных желез. Из этих отверстий животное при раздражении выпрыскивает на далекое расстояние (15 см) струйку слизи— единственное средство защиты первичнотрахейных.
Ходные конечности состоят из конического бугорка, или ножки, и сидящей на ней более короткой лапки с двумя хитиновыми коготками (•рис. 419). По степени расчлененности конечности не сложнее парапо-дий кольчецов. Движутся первичнотрахейные медленно и неповоротливо.
Покровы первичнотрахейных состоят из тончайшей, нежной, покрытой шипиками хитиновой кутикулы и из однослойного кожного эпителия. Под эпителием лежит слой соединительной ткани, а еще глубже помещается отлично развитый кожно-мускульный мешок из гладких мышц. Он состоит из наружного слоя кольцевых и внутреннего — продольных мышц (см. рис. 422). Как расположение, так и гистологическое строение мышц напоминает кольчецов.
Полость тела первичнотрахейных в общем сходна с таковой членистоногих (с. 291).
У зародышей образуются парные метамерные целомические мешки, которые позднее разрушаются, так что вторичная полость тела смешивается с первичной. Из клеточного материала целомических мешков формируются мускулатура, соединительная ткань и т. д. Сохраняются лишь небольшие участки мешков, превращающиеся в пузырьки выделительных органов и дающие стенку гонад (см. рис. 423). Однако кроме этих остатков целома сохраняется также перитонеальный эпителий на поверхности внутренних органов (кишечника и др.).
Пищеварительный канал (рис. 420) состоит из передней, средней и
задней кишок. Рот лежит на брюшной стороне головы не прямо на по
верхности тела, а на дне особой ротовой полости, образующейся посред
ством впячивания покровов головы. В ротовую полость впадает пара
длинных трубковидных слюнных желез. Затем следует глотка, а за ней
короткий пищевод, связанный с прямой длинной средней кишкой. Ко
роткая задняя кишка заканчивается порошицей на заднем конце тела.
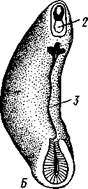
Нервная система устроена примитивно (рис. 420), но мозг состоит из
трех отделов, как у членистоногих (с. 292). Передний отдел мозга —
протоцеребрум — иннервирует глаза, второй отдел — дейтоцеребрум —
антенны, а третий отдел — тритоцеребрум — посылает нервы к передней
кишке. Нервы к челюстям отходят от основания окологлоточных кон-
нективов, а нервы ротовых сосочков — несколько
сзади. Окологлоточными коннективами мозг сое
диняется с двумя широко расставленными брюшны-
-2 ми нервными стволами. Нервные клетки равномер
но распределяются по поверхности последних и не
образуют ганглиозных скоплений. Между стволами
|
| 17 |
| Рис. 4!9. Еире- ripatus weldoni с брюшной сто роны (по Бувье): 1 — антенна. 2 — околоротовой сосочек. 3 — рот с челюстями, t — брюшные оргАны, 5 — отверстия цс-ломодуктов, 6 — ножка, 7 — половое отверстие, 8 — анус |
Рис. 420. Анатомия самки Peri-
patoides novae-zcalandiae (no
Снодграссу):
Антенна, 2 — глаза, 3 — мозг, 4 — брюшной нервный ствол, 5 — кишка, 6 — матка, 7 — яичник, 8 — задняя кишка, 9— анус, 10, 11 — яйцевод, его средняя часть, содержащая зародышей, служит маткой (6), 12 — слизистая железа, 13 — слюнная железа, 14 — проток слизистой железы, 15 — пищевод, 16 — глотка, 17 — околоротовой сосочек
имеются многочисленные тонкие комиссуры. На заднем конце тела, над задней кишкой, стволы соединяются. Итак, брюшная нервная система представляет род очень просто устроенной нервной лестницы. На всем протяжении нервных стволов от них направляются многочисленные нервы к ножкам и к стенкам тела.
|
Органы чувств. Кроме глаз и антенн имеются многочисленные кожные осязательные сосочки. Глаза лежат непосредственно над мозгом; они одиночные, построены по типу глазного пузырька и похожи на глаза полихет.
Органы дыхания представлены просто устроенными трахеями (рис. 421). На поверхности тела рассеяны многочисленные мелкие дыхальца, или стигмы. Каждое ведет в пучок длинных неветвящихся хитиновых трахейных трубочек, слепо замкнутых на концах. По ним воздух свободно проникает в глубь тела, кислород воздуха диффундирует в миксоцель, окисляя гемолим- . фу, заполняющую полость тела. Простое строение трахей привело к гипотезе о происхождении их от сильно развившихся, изменивших свою функцию кожных желез.
Кровеносная система. Сердце первично-трахейных длинное, трубчатое, тянется над кишкой от первого до предпоследнего туловищного сегмента (рис. 422, 423). Оно снабжено парными метамерными остиями и лежит, как и у Arthropoda, в перикардиальяом синусе, отделенном от остальной полости тела тонкой диафрагмой. Периферические сосуды отсутствуют.
| Рис. 421. Разрез через стигму и пучок трахей Peripatopsis capensis (из Ланга) |
Выделительная система представлена метамерно расположенными парными сег-ментарными органами, наружные отверстия которых лежат у основания конечностей. Каждый орган состоит из узкого выводного канала, переходящего потом в расширение или мочевой пузырь. Обе эти части лежат в основании конечности. От мочевого пузыря внутрь отходит извитой канал, имеющий на
внутреннем конце широкую воронку. Воронка выделительного канала открывается в небольшой целомический мешочек — остаток целомиче-ских мешков зародыша. Часть петлеобразного канала покрыта мерцательным эпителием. Общее расположение, отношение к целому (рис.423), гистологическое строение (мерцательный эпителий) и развитие (происходят из мезодермы) позволяют установить тесную связь этих органов с целомодуктами кольчецов. Интересно, что, судя по эмбриологическим данным, «слюнные» железы первичнотрахейных также представляют пару видоизмененных целомодуктов.
Половая система. Protracheata раздельнополы, причем самцы нередко значительно мельче самок. Половые железы парны, протоки же обнаруживают слияние в одном или даже двух участках на всем протяже-.нии. Пара семенников ведет в два семяпровода, образующих в начальной
части вздутия — семенные пузырьки. Затем семяпроводы сливаются в непарный семяизвергательный проток, открывающийся наружу между последней парой ног, т. е. на предпоследнем членике тела. У самки (см. рис. 420) имеются два яичника, от которых отходят яйцеводы, расширяющиеся в трубчатые матки. Последние сливаются1 в непарное
|
| 11 |
Рис. 422. Поперечный разрез через туловище Peripatoides по-vae-zealandiae (из Аверинцева):
/ — кутикула, 2 — кольцевая мускулатура. 3 — косая мускулатура, 4 и
13 — продольная мускулатура, 5 — слюнная железа, 6 — поперечная
мышца, 7 — деломодукт. * — нервный ствол, 9 — мускулатура ножки,
10 — коготки, 11 — лапка, 12 — средняя кишка. 14 — выделительное от
верстие, 15—проток слизистой железы, 16 — диафрагма, отделяющая
околосердечный участок полости тела, 17 — сердце. 18— средний отдел
полости тела (миксоцеля), 19 — боковой отдел полости тела (миксо-
• целя)
влагалище, открывающееся наружу между основаниями предпоследней пары ножек. В том сегменте, где лежит половое отверстие, отсутствуют выделительные органы. Это обстоятельство и история развития половых протоков показывают, что последние представляют пару видоизмененных целомодуктов.
Развитие.Оплодотворение внутреннее; сперматозоиды в половом протоке самца склеиваются в сперматофоры, которые или прикрепляются самцом к поверхности тела самки, или откладываются на почву. В первом случае из сперматофоров спермин проникают через трещины кожи в полость тела самки и оплодотворяют яйца, находящиеся еще в яичниках. Во втором случае самка подбирает сперматофоры с почвы краями своего полового отверстия. Почти все первичнотрахейные, кроме одного вида, живородящи.
У некоторых родов зародыши, развивающиеся в матках, срастаются в определенном участке тела со стенкой матки, развивается нечто вроде детского места, или плаценты, млекопитающих. Через плаценту и совершается питание зародыша за счет материнского организма. Развитие прямое.
Экология и распространение.Первичнотрахейные встречаются главным образом в подстилке влажных тропических лесов, под упавшими стволами деревьев, камнями. Географическое распространение их очень своеобразно, близкородственные формы обитают на разных конгинен-
|
|
| 4 А |
тах. Первичнотрахейные известны из Центральной и Южной Америки, Центральной и Южной Африки, Индии, Малайского архипелага, Новой Гвинеи, Новой Зеландии и Австралии. Наиболее характерные представители: Peripatus в Америке, к этому роду относится P. torquatus — самый крупный вид, достигающий в длину 15 см, Peripatopsis— в Южной Африке, Peripatoi-des — в Южной Австралии.
Филогения.Ряд признаков первичнотрахейных указывает на их родство с аннелидами, в частности с кл. Polychaeta. Таковы: гомономность сегментации туловища, строение конечностей, близких к параподиям, развитие целомодуктов почти во всех сегментах, наличие кож-но-мускульного мешка и гладкая мускулатура и, наконец, «пузыревидные» глаза. Вместе с тем Первичнотрахейные обнаруживают много особенностей организации, общих с членистоногими: миксоцель, превращение конечностей в ротовые органы, сердце с остиями, трахеи, общее строение мозга и характер движения: первич-нотрахейные ползают, перебирая ногами, а не посредством перистальтики кожно-мускуль-ного мешка.
Некоторые черты строения, не склоняясь ни в ту, ни в другую сторону, указывают на очень большую примитивность первичнотрахейных. Такова брюшная часть нервной системы. С другой стороны, в строении их половой системы и в эмбриональном развитии имеются явные признаки вторичной специализации.
Общие с членистоногими черты строения недостаточны для включения онихофор в состав типа Arthropoda. У первичнотрахейных нет столь характерного для последних
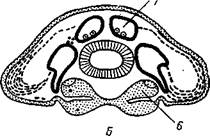
Рис. 423. Развитие целома у Peripatus. А — поперечный разрез зародыша; цело-мические мешки подразделены на 3 отдела: спинной, брюшной и боковой; 6 и В — то же, превращение спинных отделов в половые железы и образование целомодуктов за счет брюшного и отчасти бокового отделов. Стенка значительной части боковых отделов разрушается, и на их месте образуется смешанная полость тела (миксоцель) (из Давыдова):
/— спинной отдел целома, 2 — боковой, 3 — брюшной, 4 — брюшной орган, 5 — кишка, 6 — нервный ствол, 7 — половая железа, 8 — сердце, 9 — концевой пузырек целомодукта
прочного наружного скелета, хотя их тонкая кутикула и содержит хитин.
В состав головы у них входит всего три сегмента, а у членистоногих не менее четырех (с. 291). Конечности онихофор несравнимы с ногами членистоногих.
Анализ всех признаков типа Onychophora показывает, что их предками были многощетинковые кольчецы и их эволюция отчасти проходила параллельно эволюционному развитию членистоногих. В настоящее время известны ископаемые формы из кембрийских отложений, напоминающие современных онихофор, но обитавшие в море.
ТИП МОЛЛЮСКИ (MOLLUSCA)
Моллюски, или мягкотелые, образуют ясно ограниченный тип животных, ведущий начало от кольчатых червей. К моллюскам относятся главным образом водные, реже наземные животные, характеризующиеся следующими признаками.
1. Моллюски — билатерально-симметричные животные, однако у части моллюсков вследствие своеобразного смещения органов тело становится асимметричным.
2. Тело моллюсков несегментированное, лишь у ряда низших представителей обнаруживаются некоторые признаки метамерии.
3. Моллюски — вторичнополостные животные с неметамерным остаточным целомом, представленным у большинства форм околосердечной сумкой (перикардием) и полостью гонад. Все промежутки между органами заполнены соединительной тканью.
4. Тело моллюсков, как правило, состоит из трех отделов — головы, туловища и ноги. Очень часто туловище разрастается на спинную сторону в виде внутренностного мешка. Нога — мускулистый непарный вырост брюшной стенки тела, служит для движения.
5. Основание туловища окружено большой кожной складкой — мантией. Между мантией и телом находится мантийная полость, в которой лежат жабры, некоторые органы чувств и открываются отверстия задней кишки, почек и полового аппарата. Все эти образования вместе с почками и сердцем (расположенными в близком соседстве с мантийной полостью) называются мантийным комплексом органов.
6. На спинной стороне тела, как правило, имеется выделяемая мантией защитная раковина, чаще цельная, реже двустворчатая, или состоящая из нескольких пластинок.
7. Для большинства моллюсков характерно присутствие в глотке особого аппарата для размельчения пищи — терки (радулы).
8. Кровеносная система характеризуется наличием сердца, состоящего из желудочка и предсердий; она не замкнута, т. е. часть своего пути кровь проходит по системе не оформленных в сосуды лакун и синусов. Органы дыхания обычно представлены первичными жабрами — ктени-диями. Последние, однако, у ряда форм исчезают или замещаются органами дыхания иного происхождения (с. 459).
Для выделения служат почки — видоизмененные целомодукты, сообщающиеся внутренними концами с околосердечной сумкой.
9. Нервная система у примитивных форм состоит из окологлоточного кольца и четырех продольных стволов; у высших форм на стволах в результате концентрации нервных клеток формируется несколько пар ганглиев. Нервная система такого типа называется разбросанно-узловой.
10. Развитие моллюсков очень похоже на таковое многощетинковых червей; у большинства дробление спирального типа, детерминированное. У низших представителей из яйца выходит трохофора, у большинства
остальных — видоизмененная трохофорная личинка — парусник (вели-гер).
Тип Mollusca включает около 130000 видов и подразделяется на два подтипа: боконервные (Amphineura) и раковинные (Conchifera).
ПОДТИП БОКОНЕРВНЫЕ (AMPHINEURA)
Примитивные моллюски с шиповатой кутикулой, часто также с 8 ме-тамерными пластинками раковины на спинной стороне тела. Внутренностный мешок отсутствует. Нервная система с двумя парами продольных стволов, причем боковые (плевро-висцеральные) стволы переходят друг в друга позади анального отверстия. Голова без глаз и щупалец. Стато-цистов нет.
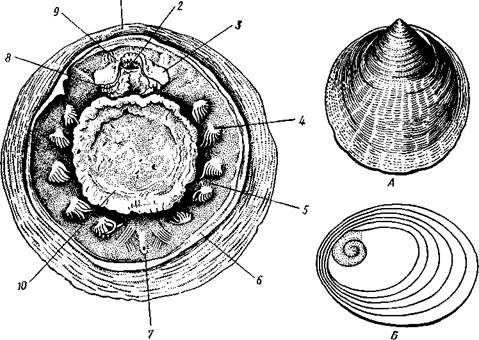
КЛАСС I. ПАНЦИРНЫЕ, ИЛИ ХИТОНЫ (LORICATA, ИЛИ POLYPLACOPHORA)
Объединяет значительное число представителей подтипа Amphineura (1000 видов). Панцирные живут главным образом в полосе прибоя, где медленно ползают по камням или прочно присасываются к ним подошвой ноги, будучи вполне защищены покрывающими их сверху пластинками раковины.
|
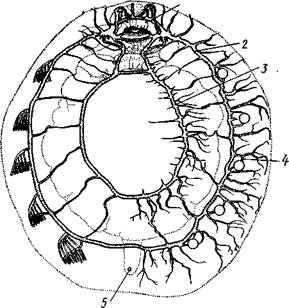
Класс Loricata характеризуется следующими особенностями. Внутренностный мешок отсутствует. Раковина, как правило, состоит из8пла-сгинок. Мантия в виде кольцевой складки равномерно окружает тело со всех сторон, прикрывая не только ногу, но и голову. Мантийная полость имеет вид круговой борозды, в которой расположены многочисленные парные ктенидии. Нервная система слагается из окологлоточного церебрального кольца и двух пар продольных нервных стволов. Головные органы чувств отсутствуют. Характерно развитие спинных органов чувств на раковине (эстеты, глаза). Панцирные — раздельнополые животные; в развитии имеется личинка — трохофора.
| Рис. 424. Хитон Tonicclla marmorea со спинной стороны (по Иванову): 1—8 — пластинки раковины, 9 — край ман- |
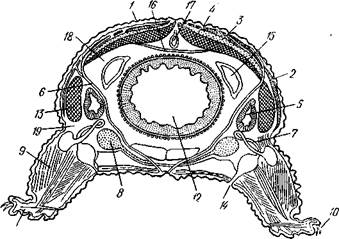
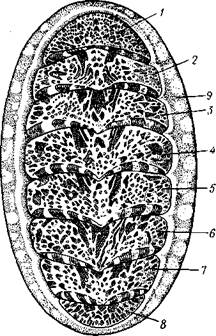
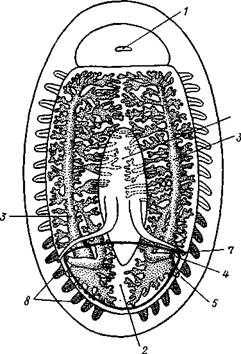
Строение и физиология.Внешняя морфология. Форма тела большей частью продолговато-овальная, несколько сплющенная в спинно-брюшном направлении (рис. 424). Размеры хитонов варьируют от нескольких миллиметров до 33 см (у толстого мясистого Cryptochiton из дальневосточных морей). Тело состоит из трех характерных для моллюсков отделов: головы, туловища и ноги. Голова слабо обособлена от туловища и лишена органов чувств; она обращена на брюшную стррону тела (рис. 425). Остальную часть брюшной поверхности занимает широкая плоская мускулистая
нога, в результате сокращения мышц которой и происходит медленное ползание моллюска по субстрату. Спинные покровы туловища хитонов образуют по всей периферии мясистую складку, нависающую со всех сторон книзу, а спереди покрывающую также голову и называемую мантийной складкой. Между этой складкой, с одной стороны, и головой и туловищем, с другой — получается промежуток в виде глубокого желобка — мантийная борозда. В полости помещаются органы дыхания и открываются также выводные отверстия пищеварительной, половой и выделительной систем.
Раковина. Спинная сторона тела покрыта раковиной, которая представляет собой продукт выделения наружного эпителия. Восемь пластинок раковины расположены друг за другом в один ряд и подвижно сочленены (см. рис. 424). Они черепицеобразно прикрывают друг друга так, что задний край первой пластинки находит на передний край второй и т. д. Вследствие такого расположения пластинок хитоны могут сворачиваться на брюшную сторону, как мокрицы и трилобиты. Пластинки
покрывают всю спинную
|
7 поверхность, оставляя об
наженной только узкую
краевую зону мантии. Ра
ковина в пределах класса
может подвергаться час
тичной редукции. Именно
пластинки раковины мо
гут уменьшаться в разме
рах (Cryptoplax) или об-
растаются толстым слоем
кожи, так что раковина
становится внутренней
(Cryptochyton).
| W // |
| Рис. 425. Хитон Tonicella marmorea, вскрытый с брюшной стороны. Большая часть органов удалена (по Иванову): / — рот. 2 — голова, 3—перерезанные кровеносные синусы (оформленные лакуны). 4 — нога. 5, 8 — ножные мышцы, 6 — правая почка, 7 — левая почка, 9 — проток почки, 10 — отверстие почки в перикардий, // — анальное отверстие, 12 — oi-фрадий, 13 — край мантии, 14 — кишка, /5 — жабры, 16 — желудочек сердца, по бокам от него два предсердия, Г/— наружное почечное отверстие. 18—наружное половое отверстие, 19 — перикардий, 20 — мантия, 21 — мышцы, связанные с раковиной, 22 — аорта |
Каждая раковинная пластинка состоит из нескольких слоев, отличающихся по составу. В наружном слое преобладает органическое рогоподобное вещество, в самом внутреннем •.— известь. Строение раковины может упрощаться за счет выпадения поверхностных слоев, что особенно часто наблюдается у видов, обладающих внутренней раковиной. В эпителии краевой зоны спины развиваются многочисленные известковые чешуйки и иглы. Каждая игла формируется одной клеткой эпителия. Снаружи эпителий одет эластичной кутикулой.
Строение раковины наложило отпечаток на
особенности строения мускулатуры Loricata. Группы мышечных пучков, прикрепляющиеся к раковинным пластинкам, следуют упорядочение одна за другой, чем напоминают метамерное расположение органов у членистых животных (в первую очередь, кольчатых червей).
|
| 12 |
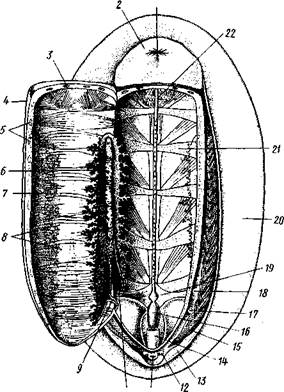
Пищеварительная система (рис. 426). Рот лежит на нижней стороне головы и ведет сначала в ротовую полость, затем в мускулистое расширение передней кишки — глотку. Со дна глотки в нее вдается продольный мускулистый валик — язык. Поверхность языка одета довольно толстой роговой кутикулой, на которой сидят в несколько поперечных и продольных рядов роговые зубцы. Это перетирающий пищу аппарат — терка, или радула. Передний конец языка свободно торчит в полость глотки, тогда как задний погружен в глубину узкого и длинного кармашка, образованного выпячиванием глоточной полости. Роговая пластинка, покрывающая язык, доходит до самого конца этого влагалища радулы. Передний край радулы от употребления постепенно стирается, но на дне влагалища имеется несколько рядов эпителиальных клеток (одонтобластов), выделяющих новые роговые зубчики. Они выдвигаются постепенно из влагалища на поверхность языка и заменяют стершиеся части радулы. Со спинной стороны в глотку открывается пара небольших слюнных желез, а несколько далее, на месте перехода глотки в пищевод, пара особых сахарных желез, секрет которых способствует превращению крахмала в сахар.
За глоткой следует более узкий пищевод, переходящий в эн-тодермальную среднюю кишку. Начальный отдел последней образует мешковидное расшире- 4-ние — желудок. В него открываются протоки большой двухло-
правая лопасть 5"
пастнои печени,
которой меньше левой и лежит несколько впереди нее. Остальная часть средней кишки, или тонкая кишка, отличается большой длиной (в несколько раз длиннее тела); она образует многочисленные изгибы, затем направляется назад и открывается при помощи короткой задней кишки на заднем конце туловища в мантийную полость. Пищей панцирным служат преимущественно водоросли, которых они соскребают при помощи радулы с камней и скал.
| Рис. 426. Хитон Tonicella marmorea, вскрытый со спинной стороны; аорта, сердце, половая система, лопасти печени удалены (по Иванову): / — слюнная железа, 2 —глотка, 3 — пищевод, 4 — желудок, 5 — отверстия печени в желудок, 6 — кишка, 7 — задняя кишка, 8 — ножные мышцы, 9 — выросты почки, 10 — внутренностная артерия, // — радула, 12 — сахарная железа, 13 — мышцы глотки |
Нервная система (рис. 427) носит примитивный характер. Она состоит из окологлоточного нервного кольца, верхняя половина которого соответствует парным мозговым, или церебральным, узлам других моллюсков, и
| I |
издвух педальных и двух плевровисцеральных нервных стволов. Педальные стволы расположены ниже и кнутри от плевровисцеральных; они проходят внутри ноги и связаны между собой многочисленными, расположенными без особого порядка поперечными перемычками. Плевровисцеральные стволы проходят под мантийным желобком и сзади, над порошицей, переходят один в другой. Кроме того, плевро-висцеральный ствол каждой стороны соединяется с педальным той же стороны множеством поперечных перемычек. В общем, нервная система получает вид как бы двойной веревочной лестницы, несколько напоминая таковую некоторых Polychaeta. Окологлоточное кольцо снабжает нервами голову, педальные стволы — ногу, а Плевровисцеральные — все остальное тело.
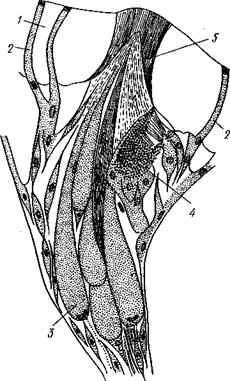
Органы чувств развиты слабо. В мантийной полости с каждой стороны залегает валик чувствительного эпителия, проходящий у основания жабр. Эти валики считаются органами химического чувства. Настоящие органы химического чувства,— осфрадии, свойственные моллюскам других групп, у хитонов чаще всего рудиментарны. Они представляют группы высоких пигментированных клеток, расположенных у основания самой крупной пары жабр (см. ниже).
|
Очень распространены у Loricata мелкие органы чувств на спинной стороне тела, или эстеты. Это эпителиальные сосочки, входящие от кожного эпителия спинной стороны в наружный слой раковинных пластинок; они содержат группу продолговатых чувствительных клеток, над которой поверхностная
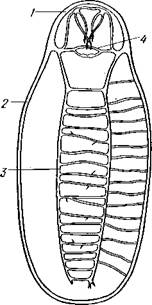
Рис. 427. Нервная система хитона Acanthochiton disorepans
(по Пельзнеру): / — церебральная нервная дуга, 2 — плевровисцеральный ствол, 3—педальный ствол, 4 — подглоточная дуга
Рис. 428. Разрез через спинной орган чувств хитона Callochiton (из Гаймен):
1 — наружный слой раковины, 2 — мик-
рэстеты, 3 — железистые клетки, 4 —
глазок, 5 — мегалэстет
пленка органического слоя раковины образует утолщение в виде рогового колпачка. Внутренние концы чувствительных клеток эстета продолжаются в нервные волокна, которые вступают в связь с плевровисцеральными стволами. По размерам различают крупные, состоящие из нескольких, а иногда даже из многих клеток мегал-эстеты, и мелкие, содержащие одну чувствительную клетку, микрэстеты. Эстеты рассеяны в большом числе на спине животного, образуя иногда более или менее правильные ряды; это органы осязания. У некоторых Loricata часть эстетов модифицируется в своеобразно устроенные глазки (рис. 428).
Настоящие глаза, органы равновесия — статоцисты и головные щупальца— у панцирных моллюсков отсутствуют.
Органы дыхания. Панцирные моллюски обладают жабрами. Число их в отличие от большинства моллюсков колеблется от 4 до 80 пар. Жабры располагаются в один ряд на каждой стороне тела в мантийной борозде (см. рис. 425), причем одна пара, лежащая позади почечных отверстий, больше других. Каждая жабра имеет вид ланцетовидной пластинки с широким основанием; на обеих плоских сторонах пластинки перпендикулярно к ней отходит по ряду лепестков, прилегающих друг к другу, как страницы книги. Таким образом, жабра имеет двоякоперистое строение. Вся поверхность ее одета мерцательным эпителием. Вдоль узкой стороны жаберной пластинки, обращенной к ноге, от основания к свободному концу жабры проходит приносящий жаберный сосуд с венозной кровью, а вдоль стороны, обращенной к мантии, от конца жабры к ее основанию идет выносящий сосуд с окисленной кровью. Веточки обоих сосудов продолжаются в жаберные лепестки. Благодаря работе ресничек эпителия вокруг жабры циркулирует вода, что способствует газообмену между кровью жаберных сосудов и внешней средой.
Кровеносная система состоит из сердца и кровеносных сосудов. Сердце лежит над кишечником в задней части тела и образовано срединным желудочком и двумя боковыми предсердиями (см. рис. 425). Кровь поступает из предсердий в желудочек через одну-две пары атрио-вентри-кулярных отверстий с клапанами, которые мешают крови возвращаться в предсердия при сокращениях желудочка. Оба предсердия сообщаются друг с другом позади желудочка. Желудочек сзади замкнут слепо, а на переднем конце продолжается в главный сосуд тела — аорту. Аорта направляется вперед, посылая от себя побочные артерии к половой железе. Из артерий кровь, постепенно отдавая кислород тканям, поступает в систему лакун, т. е. в неправильные промежутки между тканями и органами. Из лакун кровь, лишившись кислорода, собирается в два приносящих жаберных сосуда, располагающихся по бокам тела под плевровисцеральными стволами. От них ответвляются более тонкие сосуды, которые в жабрах распадаются на капилляры, где кровь окисляется и затем из каждой жабры по выносящему жаберному сосуду попадает в 2 более мощные жаберные вены. Последние идут параллельно приносящим сосудам, но только над плевровисцеральными стволами; обе жаберные вены открываются в предсердия.
Сердце окружено особым участком вторичной полости тела — околосердечной сумкой, или перикардием. Итак, кровеносная система панцирных незамкнута, но строение сердца более сложное, чем у членистоногих.
Органы выделения. Кровь освобождается от накапливающихся в ней азотистых продуктов обмена веществ, отдавая их выделительным органам— почкам. С обеих сторон кишечника залегает по V-образно изогнутому каналу, их вершины обращены вперед (рис. 429). Внутренняя,
|
обращенная к срединной линии ветвь этого канала, сообщается при помощи мерцательной воронки с перикардием, наружная открывается выделительной порой сбоку от порошицы в мантийный желобок, т. е. наружу. На всем протяжении канала от него отходят тонкие ветвящиеся слепые выпячивания. По мезодермальному происхождению и присутствию на внутреннем конце канала мерцательной воронки, открывающейся в целом (перикардий), . выделительные органы моллюсков соответствуют целомодук-там (половым воронкам) высших червей.
Половая система. Панцир-
|
| 12 Л |
1 2 3
Рис. 429. Схема выделительной и половой системы Loricata (по
Геллеру):
1 — рот, 2 — перикардий, 3—почки, 4 — внутренние отверстия почек в перикардий, 5 — наружные отверстия почек, 6 — половая железа, 7 — половые отверстия, 8 — жабры
Рис. 430. Схема поперечного разреза хитона (из Лемке и Вингстранда):
/ — яичник, 2 — аорта, 3 — продольные мышцы, 4 — раковина, 5 — эстеты, 6 — почки, 7 — плевровисцеральный ствол, * — кутикулярные шипы мантии, 9 — жабра, 10 — педальный ствол, 11 — кровеносные лакуны, 12 — кишка, \3 — приносящий жаберный сосуд, 14 — выносящий жаберный сосуд
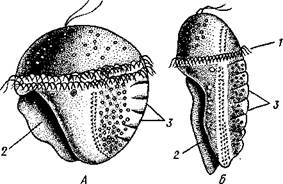
Рис. 431. Личинка хитона Ischnochiton magdaten-
sis и ее метаморфоз (по Хитсу). А — трохофора в
начале метаморфоза; Б — несколько более поздняя
стадия:
1 — ирототрох, 2 — зачаток ноги, 3 — закладка пластинок раковины
ные раздельнополы, причем их половые железы в большинстве случаев сливаются в непарный семенник или яичник, лежащий над кишечником (рис. 430). Гонада не имеет сообщения с перикардием, но снабжена двумя специальными половыми протоками, открывающимися в мантийный желобок (см. рис. 429). Органов совокупления нет, половые продукты выводятся прямо в воду.
Развитие.Из яйца хитонов развивается личин-
ка, снабженная предротовым венчиком ресничек, теменной пластинкой с султаном длинных ресничек и очень похожая на трохофору кольчецов. Затем она приобретает некоторые признаки, характерные для панцирных моллюсков: на спинной стороне закладываются особые ямки, в которых формируются раковинные пластинки; на брюшной стороне появляется зачаток ноги в виде выступа, покрытого ресничками, и зачатки двух пар небольших глазков (рис. 431). Личинка сначала ведет плавающий образ жизни, потом, по мере дальнейшего развития органов, свойственных взрослым хитонам, личиночные признаки — теменной султан, венчик ресниц, глаза — исчезают, и молодое животное опускается на дно.
КЛАСС II. БОРОЗДЧАТОБРЮХИЕ, ИЛИ БЕСПАНЦИРНЫЕ (SOLENOGASTRES, ИЛИ APLACOPHORA)
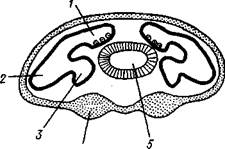
Класс объединяет Amphineura, лишенных раковины и ноги. Червеобразное тело покрыто кутикулой, несущей многочисленные известковые шипы. Мантийная полость находится на заднем конце тела.
|
|
| Рис. 432. Представители бороздчатобрюхих моллюсков. А — Chaetoderma nitidulum; Б — Neomenia carinata (из Гаймен); 1 — жабры. 2 — ротовое отверстие, 3 — брюшная бороздка |
Бороздчатобрюхие обитают в илу или поселяются на колониях гидроидных полипов, гидрантов которых объедают. Это морские малоподвижные животные, встречающиеся преимущественно на глубинах. Число известных видов около 150. Их длинное червеобразное тело иногда достигает 30 см. На месте ноги у части бороздчатобрюхих имеется продольный мерцательный желобок с узким килем, представляющим рудимент ноги. Радула рудиментарна или отсутствует. Кишка прямая, многие виды лишены желудка и печени. Жабры отсутствуют (Рагате-nia, Neomenia; рис. 432,5) или же они представлены одной парой, расположенной на заднем конце тела (Chaetoderma; рис. 432,Л). Нервная система очень близка к таковой Loricata. Среди бороздчатобрюхих встречаются как гермафродитные,
так и раздельнополые формы. Развитие осуществляется с метаморфозом, во время которого у животного на одной из стадий закладываются спинные пластинки, позднее отпадающие и заменяющиеся мелкими спи-кулами.
ПОДТИП РАКОВИННЫЕ (CONCHIFERA)
Моллюски с известковой раковиной, цельной или разделенной на две боковые створки. Покровы без кутикулы. Внутренностный мешок часто хорошо развит. Нервная система чаще разбросанно-узлового типа. Боковые нервные стволы сзади соединяются под задней кишкой. На голове расположены глаза и щупальца. Имеются статоцисты.
| 15—763 |
Различаются 5 классов: Моноплакофоры (Monoplacophora), Брюхоногие (Gastropoda), Лопатоногие (Scaphopoda), Пластинчатожаберные (Lamellibranchia) и Головоногие (Cephalopoda).
КЛАСС I. МОНОПЛАКОФОРЫ (MONOPLACOPHORA)
В ископаемом состоянии эти моллюски известны уже давно из отложений кембрия, силура и девона. Они обладали колпачковидной, блюд-девидной или спирально закрученной раковиной. По внешнему сходству с брюхоногими моллюсками они долго причислялись к классу Gastropoda и лишь в 1940 г. были выделены в особый класс (с. 500).
В 1952 г. неожиданно был найден современный представитель класса — Neopilina galatheae. Эта замечательная находка была сделана в восточной части Тихого океана на глубине 3590 м датской зоологической морской экспедицией на судне «Галатея».
Тело Neopilina состоит из небольшой головы, довольно высокого туловища и дисковидной ноги (рис. 433). Голова, лежащая спереди на брюшной стороне тела, почти не обособлена от туловища, несет ротовое отверстие. Перед ним располагается пара щупалец и особая кожная складка — велум. Глаза отсутствуют. У концов велума около переднего края ноги лежит пара кустикообразных щупалец, вероятно, имеющих обонятельную функцию. Туловище N. galatheae покрыто низкой конусовидной раковиной с круглым нижним краем и вершиной, сдвинутой вперед (рис. 434, Л). Диаметр раковины до 3 см. Колпачковидная раковина
|
Рис. 433. Neopilina galatheae с брюшной стороны Рис. 434. Neopilina galatheae. A — ра~
(поЛемке): конина со спинной стороны; Б — вер
у—край р.чкпнины, г —рот, ,? —велум. •/ — жабра. 5— хушка раковины с личиночной рако-
мантийная борозда, 6 - край _мантии, 7 - анус. S-орган ВИНОЙ (ПО Лемке И Вингстранду)
химического чувства, 9 — голова, 10 — нога

|
| 12 |
| 11 |
современных моноплакофор, по-видимому, возникла в результате вторичного упрощения. В пользу этого предположения говорит наличие спиральных раковин у древних ископаемых форм. Кроме того, даже у N. ga-Latheae в процессе развития сначала формируется спирально закрученная раковина, сохраняющаяся и у взрослых животных на самой вершине колпачковидной раковины (рис. 434, Б). Нижние края туловища переходят в кольцевую кожную складку, окружающую голову и ногу и прилегающую к краям раковины. Это мантия, чрезвычайно похожая на таковую у панцирных. Между мантией, головой и ногой находится довольно широкая мантийная борозда. Нога, расположенная позади головы, имеет широкую плоскую подошву. Мускулатура Neopilina обнаруживает отчетливо выраженную метамерию — име-ется 8 пар мышц, идущих от ноги к спинной стороне раковины (рис. 435). Позади ноги лежит небольшой сосочек с анальным отверстием, а по бокам от йоги на дне мантийной борозды располагаются 5—6 пар перистых жабр.
| Рис. 435. Схема организации Neopilina (по Лемкс и Вингстранду): / — голошюе щупальце, 2 — ножные мышцы, 3 — почка, 4 — наружное почечное отверстие, 5 — жабра, 6—желудочек сердца, 7 — анальное отверстие, 8 — предсердие, 9—половая железа, 10 — проток, соединяющий почку с целомом, // — плевровисцеральиый ствол, 12 — велум |
Пищеварительная система состоит из глотки, пищевода, желудка, средней и задней кишки. Имеется хорошо развитая радула с многочисленными роговыми зубами. В желудок двумя самостоятельными отверстиями открывается пара симметричных лопастей печени, а в его полости находится хрустальный столбик (с. 457). Длинная средняя кишка образует несколько больших петель и заканчивается довольно широкой задней кишкой.
|
| Рис. 436. Схема поперечного разреза Neopilina (из Лемке и Вингстранда): |
| — аорта, 2 — дорзальный целом, 3 — раковина, 4—поч-а, 5 — плевровисцеральпый стиол, 6 — жабра. 7 — нога, — педальный ствол, 9 — половая железа, 10 — кровенос- мяа nawuzt II — иы[[||^э |
| ная лакуна, II — кишка |
Кровеносная система образована сердцем, состоящим из пары желудочков и двух пар предсердий, и кровеносными сосудами. От желудочков отходит аорта (рис. 435), по которой кровь поступает в систему лаку-нарных пространств, окружающих кишечник, гонады, печень и другие внутренностные органы. Венозная кровь из лакун поступает в органы дыхания — жабры. Окисленная кровь из задней пары жабр изливается в
| 1Я* |
заднюю пару предсердий, кровь из остальных ктенидиев поступает в продольные боковые синусы, сообщающиеся с передней парой предсердий. Из предсердий кровь направляется в желудочки, расположенные по сторонам кишечника. Передние оттянутые концы желудочков объединяются и образуют аорту.
Каждый желудочек с прилегающими предсердиями заключен в тонкостенный целомический мешок — перикардий. Кроме перикардиальных
|
целомов имеются очень
./ обширные парные дор-
зальные целомы (рис.
436), сообщающиеся
Дата добавления: 2015-01-24; просмотров: 2330;