Створка, вмещающая тело животного, 2 — створка, играющая роль крышечки
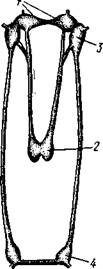
Рис. 466. Типы замков пластинчатожаберных. А — равнозу-бый замок; Б — разнозубый замок (из Ремане): 1 — главные зубы, 2 — второстепенные зубы
другой, более выпукла и вмещает все тело, тогда как правая; створка тонка и функционирует лишь в качестве крышечки. Особенно сильна разница между обеими створками у некоторых ископаемых форм (рис. 465); у рудистов, например, одна из створок имеет вид высокого конуса, а другая, по-видимому, служила крышечкой (Hip-purites). Некоторые пластинчатожаберные, например корабельный червь (Teredo; см. рис. 476), обладают сильно редуцированной раковиной, маленькие створки которой прикрывают лишь 1/20 часть тела. В типичных случаях обе створки выпуклы, причем наиболее выпуклые точки их лежат у спинного края раковины и называются макушками створок. Макушки являются наиболее старыми, начальными частями створок, к которым впоследствии присоединяются по краю все новые слои извести. Соответственно на раковинах можно различать ежегодные слои прироста, идущие параллельно свободному краю раковины и позволяющие определять возраст животного. На спинной стороне тела створки связаны между собой, во-первых, лигаментом, во-вторых, замком. Лигамент состоит из эластического вещества и соединяет обе створки в виде короткой поперечной ленты. Наружный слой лигамента непосредственно переходит в таковой створок, так что раковина, в строгом смысле слова, состоит из одного куска, перетянутого и перегнутого на спине. Вследствие своей эластичности лигамент держит обе створки полуоткрытыми.
Замком называется соединение створок при помощи зубовидных отростков (зубов) спинного края, входящих в углубления противоположной створки. Различают 2 главных типа замков: равнозубый со значительным числом зубьев одинаковой величины и формы (Nucnla; рис. 466,Л; Area) и разнозубый с небольшим числом зубов разной формы (рис. 466,Б); первый тип зубов более древний. У некоторых форм (например, беззубки — Anodonta) замок может отсутствовать и тогда створки соединены лишь лигаментом.
Для захлопывания раковины служат замыкательные мышцы, которых бывает две или одна. Они имеют вид толстых мускульных пучков, идущих поперек тела моллюска от одной створки к другой. Как в местах
|
|
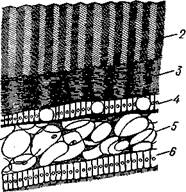
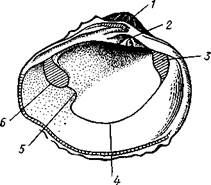
Рис. 467. Створка раковины с внутренней стороны (по Лангу):
Макушка раковины, 2 — спинной край с замком, 3 — отпечаток переднего мускула-замыкателя, 4 — отпечатки мышц, прикрепляющих край мантии к раковине (мантийная линия), 5 — «сифонный заливэ — место отхож-дения трубок сифонов, 6 — отпечаток заднего мускула-замыкателя
Рис. 468. Разрез через раковину и мантию беззубки Anodonta (по
Лейдигу):
Конхиолиновый слой, 2 —- фарфоро-видный, или призматический, слой, 3 — перламутровый слой, 4 — эпителий наружной поверхности мантии, 5 — соединительная ткань мантии, б — эпителий внутренней поверхности мантии
прикрепления мышц к створкам, так и по краю мантийных складок на внутренней поверхности раковины получаются легкие отпечатки, по присутствию или отсутствию которых можно судить о числе и расположении замыкательных мускулов, о степени развития сифонов и т. п. (рис. 467). Отпечатки хорошо сохраняются на ископаемых раковинах, что позволяет наряду с некоторыми другими данными судить о внутреннем строении давно вымерших животных.
Наружный тонкий слой раковины (рис. 468), или периостракум, состоит из органического вещества, конхиолина, и нередко стирается на выпуклых частях створки (у макушки). Под ним залегает призматический, или фарфоровидный, слой, слагающийся из тесно прилегающих друг к другу призмочек углекислой извести, поставленных перпендикулярно поверхности раковины. Этот слой обладает значительной толщиной. Самый внутренний слой, перламутровый, образуется тончайшими, лежащими в несколько слоев известковыми листочками, между которыми залегают столь же тонкие прослойки конхиолина. В перламутре имеет место интерференция световых лучей; вследствие этого перламутр блестит и переливает разными цветами. Перламутровый слой подстилается эпителием мантии, который и выделяет раковину. Этот же эпителий у некоторых Lamellibranchia формирует жемчуг. Если какие-нибудь очень мелкие частицы, например омертвевшие клетки или зернистые продукты выделения, или тельца постороннего происхождения, например песчинки, нередко паразиты, попадут в промежуток между раковиной и эпителием маитии, то они все больше и больше обволакиваются концентрическими слоями перламутра и превращаются в жемчужины.
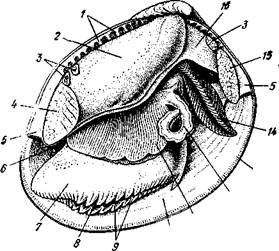
Пищеварительная система (рис. 469). Рот расположен на переднем конце тела над основанием ноги. По бокам рта имеются 2 пары длинных треугольных ротовых лопастей. Они покрыты ресничками, подго-
|
| 17 |
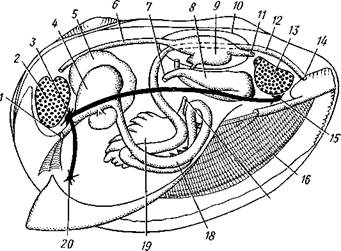
Рис. 469. Схема внутреннего строения пластинчатожаберного
моллюска (из Реманс):
Рот, 2 — передний мускул-замыкатель, 3 — церебро-плевральный ганглий, 4 —желудок, 5 — печень, 6 — передняя аорта, 7 — наружное отверстие почки, « — почка, открывающаяся в перикардий, 9 — сердце, 10—перикардий, 11 — задняя аорта, 12 — задняя кишка, 13 — задний мускул-замыкатель. 14— анальное отверстие. 15 — висцеро-париеталь-ный ганглий, 16—жабры. 17 — отверстие гонады, 18 — средняя кишка, 19 — гонада, 20 — педальный ганглий
няющими пищевые частицы к ротовому отверстию. Редукция головы вызывает атрофию тех частей кишечника, которые у прочих моллюсков помещаются в голове, а именно глотки, терки, челюстей и слюнных желез.
Рот ведет прямо в короткий пищевод, который открывается в меш-ковидный желудок. Недалеко от места впадения пищевода, но более вен-трально, от желудка отходит средняя кишка. В заднюю часть желудка открывается отверстие слепого мешковидного выроста, в полости которого формируется прозрачный студенистый стержень — кристаллический стебелек. Он состоит из мукопротеинов и ферментов (амилазы, гликоге-назы и др.). Свободным концом стебелек вдается в просвет желудка, где постепенно растворяется, высвобождая пищеварительные ферменты, которые и обеспечивают первичную обработку пищи. По бокам желудка помещается парная, хорошо развитая печень, состоящая из множества мелких долек и впадающая своими протоками в желудок.
Средняя кишка спускается от желудка в основание ноги, делает несколько изгибов и затем направляется по спинной стороне туловища к его заднему концу. Она переходит в заднюю кишку, которая обычно пронизывает желудочек сердца и заканчивается порошицей над задней замыкательной мышцей.
Будучи малоактивными, подчас и неподвижно прикрепленными (например, устрица — Ostrea, мидия — Mytilus и др.) животными, пластинчатожаберные питаются пассивно. Пищей им служат мелкие взвешенные в воде частицы — детрит, планктонные организмы и бактерии, которые моллюски отфильтровывают из воды, проходящей через мантийную полость.
|
Нервная система (рис. 470) состоит из 3 пар ганглиев. Цереброплевральные ганглии представляют продукт слияния двух пар узлов; это доказывается тем, что у примитивных Protobranchia плевральные узлы еще несколько обособлены от церебральных. Цереброплевральные узлы соединяются над глоткой тонкой церебральной комиссурой. В ноге залегает пара педальных ганглиев, которые соединяются с церебро-плевральными посредством двух длинных коннекти-вов. Еще более длинные коннективы идут от церебро-плевральных узлов к паре висцеропариетальных ганглиев, лежащих под задним мускулом-замыкателем. Эти ганглии кроме внутренностей иннервируют ос-фрадии и жабры.
Органы чувств развиты слабо, чему, по-видимому, служит причиной малоподвижный роющий образ жизни. У основания жабр имеются осфрадии, а в соседстве с педальными ганглиями всегда находятся два ста-тоциста.
| Рис. 470. Схема нервной систем и Lamellibranchia (по Гессе): 1 — церебральные ганглии, 2 — педальные ганглии, 3 — плевральный ганглий, 4 — висцерона]^е-тальный ганглий |
Головные щупальца и глаза, гомологичные соответствующим образованиям брюхоногих, отсутствуют. Имеются случаи, когда типичные ограны зрения вторично возникают или по всему краю мантии (гребешок— Pecten), пли по оторочке сифонов (сердцевидка— Cardium). У гребешка мантийный край несет свыше сотни отдельных инвертированных глаз довольно сложного строения.
Органами осязания пластинчатожаберных служат отчасти околоротовые лопасти, а также различные
щупальцевидные придатки, развивающиеся по свободному краю мантии (Pecten) или по краям сифонов.
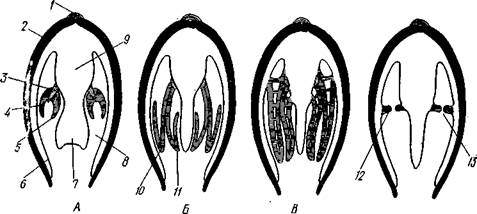
Органы дыхания (см. рис. 463) представлены рядом модификаций типичных ктенидиев. У представителей примитивного отряда Protobran-chia по бокам основания ноги в мантийной полости лежит по двояко-перистому ктенидию, состоящему из общей оси и двух рядов треуголь-
|
| Г |
Рис. 471. Жабры Lamellibranchia. Схематические поперечные разрезы через тело в области жабр. А — Protobranchia; Б — Filibranchia; В — Eulamcllibranchia; Г—Septi-
branchia (из Ланга):
/ — лигамент, 2 — створка раковины, 3 — ось ктенидия, 4 — наружный лепесток ктенидия, 5 — внутренний лепесток ктенидия, 6—мантия, 7 — нога, 8 — мантийная полость, 9 — туловище, 10—наружная нить, состоящая из нисходящего и восходящего колен, 11 — внутренняя нить, 12 — мускулистая перегородка, произошедшая за счет участков жабры, прирастающих к мантии и ноге,
13 — отверстие в перегородке
лых боковых жаберных лепестков (рис. 471). Одним краем оси каждый ктенидий прирастает к потолку мантийной полости, а концы лепестков несколько выдаются в полость мантии.
У нитежаберных (Filibranchia) лепестки обоих рядов необычайно удлиняются и принимают вид жаберных нитей, настолько длинных, что каждая нить, свешиваясь книзу, образует сначала нисходящее колено, а затем загибается кверху и дает восходящее колено. У внутреннего ряда лепестков восходящее колено направлено к ноге, у наружного — к мантийной складке (рис. 471,Б). У части Filibranchia жаберные нити свободны. У других представителей они связаны соединительноткаными мостиками. У отряда Eulamellibranchia, наконец, соединение между жаберными нитями становится еще более полным. В результате каждая половина жабры превращается в двухслойную решетчатую пластинку (рис. 471.В). Таким образом, каждая из четырех жабр Lamellibranchia на самом деле соответствует лишь одной половине настоящего ктенидия. Эпителий жабр во многих местах имеет мерцательный характер.
У небольшого отряда Septibranchia ктенидий атрофируются, а в мантийной полости образуется мускульная горизонтальная перегородка, делящая полость на два участка: нижний и верхний, или дыхательный (рис. 471,Г); в последнем и происходит газообмен.
Кровеносная система. Сердце пластинчатожаберных помещается на спинной стороне тела и лежит в тонкостенной околосердечной сумке (перикардий). Как показывает история развития, зачаток сердца закладывается парным и у некоторых низших Lamellibranchia, а именно у Area животное обладает двумя сердцами. У прочих пластинчатожабер-
ных оба зачатка сливаются и образуют непарное сердце, состоящее из желудочка и двух предсердий. У примитивных форм отряда Protobran-chia слияние происходит над кишкой. У большинства Lemellibranchia правый и левый зачатки охватывают заднюю кишку и сливаются под и над нею, следствием чего и является пронизывание желудочка задней кишкой (см. рис. 469) — черта, характерная для пластинчатожаберных.
От желудочка берут начало два мощных артериальных сосуда — передняя и задняя аорты. Передняя аорта идет над кишкой вперед и отдает от себя артерии к внутренностям, ноге и передней части мантии. Задняя аорта следует назад под кишечником и вскоре распадается на две задние мантийные артерии. Из артерий кровь попадает в систему лакун в соединительной ткани и, наконец, собирается в большую продольную венозную лакуну, лежащую под перикардием. Из лакуны кровь направляется в проходящий вдоль основания каждой жабры приносящий жаберный сосуд, проникает отсюда в жаберные нити, окисляется и возвращается в выносящий жаберный сосуд, проходящий параллельно приносящему. Выносящие жаберные сосуды сообщаются с предсердиями сердца, из которых кровь проходит в желудочек (см. рис. 469).
Выделительная система состоит из пары почек, которые лежат в задней половине тела по бокам и несколько ниже кишки. Они имеют вид двух обширных трубчатых мешков с железистыми стенками. Каждый мешок сложен по длине-вдвое так, что принимает V-образную форму с углом, обращенным назад. Обе передние ветви заканчиваются отверстиями; одним из них почка сообщается с перикардием, а другим — с мантийной полостью.
По своему происхождению почки пластинчатожаберных — типичные
целомодукты. /
Известное участие в выделении принимают и стенки перикардия. Клетки передней половины перикардия имеют железистый характер и образуют перикардиальные железы. Последние иногда обособляются от остального перикардия в виде двух мешков — кеберовых органов, сообщающихся с ним отверстиями. Продукты выделения этих желез попадают в перикардий, а оттуда выводятся через почки наружу.
|
|
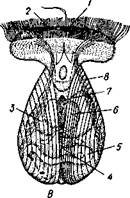
Рис.472. Развитие Dreissena polymorpha (из Мак-Брайда). А — трохофора, вид с
брюшной стороны; Б — парусник, вид спереди; В — то же, сбоку:
/ — теменная пластинка с теменным султаном ресниц, 2 — прототрох, 3 — рот, 4 — задний султан ресниц, и — нога, 6 —зачаток жабры, 7 — анальное отверстие, 8 — мускул-замыкатель, 9 — педальный ганглий, 10 — висцеральный ганглий, // — зачаток сердца, 12 — средняя кишка, 13 — створка раковины, 14 — мускульные тяжи, 15 — печень, 16 — парус
Половая система. В громадном большинстве Lamellibranchia раздельнополы. Половые железы парны и залегают в переднем отделе туловища, заходя и в основание ноги. Они имеют вид двух дольчатых, гроздевидных образований. У более примитивных Protobranchia, а также у ряда других форм (Pecten, Ostrea и др.) гонады не имеют собственных выводных протоков и открываются в почки. У большинства пластинчатожаберных, однако, дифференцируются специальные яйцеводы или семяпроводы, открывающиеся наружу по бокам основания ноги, рядом с отверстиями почек.
Развитие.Оплодотворение чаще всего наружное. Дробление идет приблизительно так же, как и у Gastropoda (с. 431), в результате получается личинка трохофорного типа (рис. 472,/4). В дальнейшем развитие интересно тем, что раковина закладывается на спине трохофоры сначала в виде цельной пластинки, которая лишь позднее перегибается по срединной линии и становится двустворчатой, причем место перегиба сохраняется в виде лигамента.
Трохофора. в результате ряда изменений превращается в характерную для многих моллюсков личинку — парусник (велигер; рис. 472). Верхняя часть трохофоры с прототрохом преобразуется в покрытый длинными ресничками диск — парус, служащий для плавания. В центре этого диска находится теменная пластинка с султаном чувствительных ресниц. Двустворчатая раковина парусника хорошо развита и покрывает все тело личинки; при плавании парус выставляется из раковины. Организация парусника во многих отношениях уже очень близка к таковой взрослого моллюска. Имеются зачаток ноги, мантия, ганглии нервной системы, желудок, печень и т. д., но органами выделения являются еще протонефридии.
После некоторого периода планктонной жизни парусник оседает на дно, причем часто прикрепляется биссусовой нитью, теряет парус и постепенно превращается в молодого моллюска.
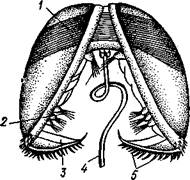
Развитие пресноводных форм (сем. Unionidae), например беззубки, обнаруживает интересные черты. Яйца откладываются в жабры (в промежуток между наружными и внутренними жаберными листками). Здесь из них развиваются двустворчатые личинки — глохидии (рис.473),
|
| Рис. 473. Глохидии беззубки Апо-donta celensis с полураскрытыми створками, вид спереди (по Гер- берсу): / — личиночный мускул-замыкатель, 5—пучки чувствительных щетинок. 3 — краевой зубец раковины, 4 — нить бис суса, 5 — зубчики на краевом зубце |
сильно отличающиеся от материнского организма. Створки глохидия округлой формы; на их брюшном крае имеется зубец с загнутыми острыми крючками. За-мыкательных мышц одна, а не две, как у взрослого животного. Большинство органов еще недоразвито: дога рудиментарна, жабр нет. У личинки посреди брюшной поверхности туловища выдается длинная липкая нить биссуса, торчащая из отверстия биссусовой железы. Когда мимо моллюска, содержащего глохидии, проплывает рыба, моллюск выталкивает через выводной сифон личинок в окружающую воду. При помощи биссусовой нити и шиповатых створок глохидии прикрепляются к жабрам или плавникам рыбы. Вокруг вызванной глохидиями маленькой ранки раздражаемый эпителий рыбы разрастается и постепенно покрывает личинку. Внутри получившейся опу-
холи глохидий питается за счет хозяина, увеличивается в размерах и в течение нескольких недель проходит дальнейшее развитие. Он постепенно превращается в миниатюрного моллюска, опухоль лопается, и молодая ракушка вываливается из нее и падает на дно. Итак, развитие Unionidae связано с временным паразитизмом, выгодным и в смысле питания, и в смысле расселения медлительных ракушек на далекие расстояния.
Экология,Пластинчатожаберные — малоактивные животные, живут на дне водоемов, нередко наполовину или целиком зарывшись в грунт. Некоторые формы ведут неподвижный образ жизни, прикрепляются к скалам или водорослям нитями биссуса (Mytilus, Pinna и др.) или прочно прирастают к субстрату створкой раковины (Ostrea, Pinctada, ископаемые рудисты; см. рис. 465, и др.). Они пропускают через мантийную полость огромные количества воды, постоянный приток которой обеспечивается работой ресничного эпителия мантии, жабр и ротовых лопастей. В тех местах, где пластинчатожаберных особенно много, например на устричных и мидиевых банках, эти моллюски становятся мощными естественными очистителями воды (биофильтраторами). Подсчитано, что ^идии, заселяющие 1 м2 дна, за сутки могут профильтровать до 280 м3 воды. В этом отношении пластинчатожаберные обнаруживают сходство с губками.
Некоторые пластинчатожаберные, особенно Pholas, Lithophaga, обладают способностью сверлить ходы в мягких горных породах, например известняках. Известно, что колонны храма Ссраписа, близ Неаполя, проточены Lithophaga до высоты человеческого роста. Присутствие Lithophaga в колоннах показывает, что со времени постройки храма (в первые века нашей эры) суша в данном районе погружалась ниже уровня моря (и в это время Lithophaga проточили ходы), а затем в результате вулканической деятельности суша опять поднялась.
Полезное и вредное значение Lamellibranchia.Некоторое полезное значение пластинчатожаберные имеют как источник перламутра и жемчуга. Для выделки перламутровых пуговиц и других изделий пригодны многие виды из пресноводного семейства Unionidae. Особенно богаты ими реки Северной Америки, где имеется несколько десятков промысловых видов этого семейства. В Европе, и в частности в СССР, количество их невелико: Unio pictorum, U. lumidus, Margaritifera и др.
Для поделок пригодны раковины с толщиной перламутрового слоя около 2,5 мм.
Образование хорошего жемчуга наблюдается у очень немногих форм. Наиболее ценится жемчуг морской жемчужницы Pinctada, встречающейся в Красном море, Индийском и Тихом океанах. Она живет на небольших глубинах (5—15 м) и раньше вылавливалась ныряльщиками. Теперь в некоторых странах (Япония) устраивают специальные хозяйства для разведения жемчужниц, которых подвергают несложной операции, стимулирующей образование жемчуга. Пресноводная жемчужница (Margaritifera), доставляющая довольно хороший жемчуг, хотя и некрупных размеров, встречается в реках и озерах нашего Севера.
Многие морские пластинчатожаберные употребляются в пищу, по-видимому, с незапамятных времен, на что указывает обнаружение раковин в «кухонных отбросах» каменного века. Наибольшую ценность в этом смысле представляют устрицы (Ostrea; рис. 474), которых еще римляне выращивали в особых бассейнах. В настоящее время большой промысел устриц имеется в Англии, Франции, США и Японии. В Англии число ежегодно потребляемых устриц превышает 2 млрд. Моллюсков не только собирают на устричных банках, т. е. на отмелях Северного моря и Атлантического океана, но и разводят. Из других съедобных моллюс-
16-763 48!
ков нужно упомянуть дальневосточного гребешка (Pecten yessoensis) и мидию (Mytilus edulis, рис. 475). Мидии встречаются у берегов, в полосе отлива. Мировая добыча мидий составляет около 2—2,5 млн. ц в год. В нашей стране тоже ведется интенсивный промысел гребешков и мидий и проводятся работы по их искусственному разведению.
Вредным представителем класса Lamellibranchia является корабельный червь, или шашень (Teredo navalis). Шашень (рис. 476) — белова-

|
|
Рис. 474. Молодые устрицы Ostrea на куске дерева (из Догеля)
Рис. 475. Мидия Mytilus edulis, прикрепленная к субстрату при помощи нитей биссуса (из Гешелера)
тое, червеобразное животное до 10 см длины с раковиной, редуцирован
ной до степени двух маленьких пластиночек. Шашень точит длинные хо
ды в деревянных днищах судов, сваях пристаней и т. п. У нас шашень
встречается в Черном море и на Дальнем Востоке. /
- Широко распространена в некоторых реках и озерах европейской части СССР и в Аральском море дрейсена — Dreissena polymorpha
(рис. 477). Моллюск прикрепляется биссусом к различным подводным предметам. В некоторых водоемах дрейсены размножаются в таких количествах, что нарушают нормальную работу различных гидротехнических сооружений и могут проникать в водопроводные трубы, которые они закупоривают, и, погибая, становятся причиной порчи питьевой воды.
Палеонтология. Ископаемые остатки пластинчатожаберных известны с кембрия. К числу наиболее древних относятся формы с равнозубым замком, несколько напоминавшие современных Nucula (рис. 478). Их считают родоначальниками многих ископаемых и современных семейств пластинчатожаберных. В ордовике число и разнообразие форм заметно возрастает, а в силурийских отложениях уже можно найти представителей многих семейств, сохранившихся и поныне (Mytilidae, Pteriidae и др.). В мезозое количество и разнообразие пластинчатожаберных резко увеличивается. Особый интерес представляют рудисты (юра — мел) с их прикрепленной неравностворчатой раковиной, иногда достигавшей 1,5 м высоты (см. рис. 465, с. 474).
У некоторых видов (Requienia) крупная нижняя створка, вмещающая тело моллюска, закручена спирально, напоминая раковину улиток. Рудисты были обитателями теплых морей, часто поселявшимися на коралловых рифах. В кайнозое достигли наибольшего расцвета формы, обладавшие разнозубым замком. Многочисленность и хорошая сохранность пластинчатожаберных позволяет их считать важными руководящими окаменелостями.
Классификация. Для пластинчатожаберных принимается несколько различных систем. Классификация этой группы может основываться на разных признаках — на особенностях строения замка, мускулов-замыкателей раковины, жабр. По последнему признаку пластинчатожаберных делят на 4 отряда.
Отряд 1. Первичножаберные (Protobranchia) — небольшая группа наиболее примитивных пластинчатожаберных, для которых характерны типичные ктенидии, нога с

|
| 13 |
| 12 |

|
| 10 |
Рис. 477. Раковина Dreissena
Рис. 476. Корабельный червь, или шашень, Teredo navalis и его ходы в куске дерева (по Мейеру и Мебиусу)
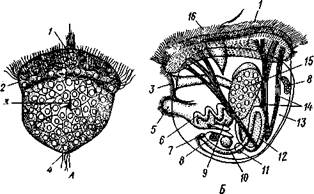
Рис. 478. Вид с левой стороны Nucula tenuis (Protobranchia). Левая створка раковины и мантия удалены (по Иванову):
7 —выросты мантии между зубцами таксодонтного валка, 2 — туловище, 3 — мускулы ноги, 4 — передний мускул-замыкатель. 5 — часть обрезанной левой мантии, 6 — ротовое отверстие, 7 — нога, 8—подошва ноги, 9 — па-пиллы ноги, 10 — правая мантия, // — ротовые лопасти. 12 — придатки ротовых лопастей, 13—мантийные мышцы, 14 — левый ктенидии, 15 —задний мускул-замыкатель, 16 — слизистая мантийная железа
подошвой, статоцист, плевральные ганглии обособленные от церебральных, отсутствие специальных половых протоков. Живут преимущественно в северных морях, обычно мелкие формы. Представители: Joldia, Nucula (рис. 478).
Отряд 2. Нитежаберные (Filibranchia). Жаберные лепестки вытянуты в длинные, сложенные пополам нити (см. рис. 471, Б). Представители: Ноев ковчег — Area noae, мидия — Mytilus (см. рис. 475); гребешок — Pecten, устрица — Ostrea (см. рис. 474); морская жемчужница — Pinctada margaritifera; Lithophaga, проделывающая ходы в известняковых породах.
Отряд 3. Настоящие пластинчатожаберные (Eulamellibranchia). Жабры превращены в двойные решетчатые пластинки. Объединяет большинство пластинчатожаберных, а именно все наши пресноводные ракушки (жемчужница — Margariiifera, Unio, Anodon-
| 16* |
ta, Dreissena) и многие морские виды (сердцевидка — Cardium; корабельный червь, или шашень — Teredo (см. рис. 476) и камнеточец Pholas способны проделывать ходы в дереве, в мягких горных породах; Tridacna — типичный обитатель коралловых рифов и др.), Отряд 4. Перегородчатожаберные (Septibranchia).К ним относятся небольшие морские, преимущественно глубоководные моллюски, с редуцированными жабрами. Их мантийная полость поделена на 2 части мускулистой поперечной перегородкой с отверстиями. Газообмен осуществляется в верхней части мантийной полости.
|
| 11. |
| 12 |
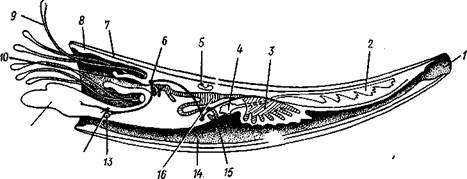
Рис. 479. Схема продольного разреза лопатоногого моллюска (из Кестнера): / — отверстие на вершине раковины и верхнее мантийное отверстие, 2 — половая железа, 3 — печень, 4 — почка, 5 — перикардий с сердцем, 6 — церебральный ганглий, 7 — раковина, 8 — мантия, 9 — ловчие щупальца, 10 — ротовое отверстие, // — нога, 12 — педальный ганглий, 13 — статоцист, 14 — мантийная полость, 15 — анальное отверстие, 16 — желудок
КЛАСС IV. ЛОПАТОНОГИЕ (SCAPHOPODA)
Этот класс объединяет небольшое количество морских моллюсков (300 видов), совмещающих некоторые черты пластинчатожаберных (Lamellibranchia) с признаками брюхоногих (Gastropoda). По внешнему виду Scaphopoda скорее напоминают брюхоногих моллюсков, обладая цельной раковиной в виде слегка изогнутой конической трубки с двумя отверстиями по концам. Из более широкого переднего отверстия высовываются голова и нога. Сходство с брюхоногими проявляется и в организации пищеварительной системы. В то же время симметричность тела, строение нервной системы, характер мантийной полости и циркуляция воды в ней приближают лопатоногих к двустворчатым моллюскам.
Лопатоногие ведут роющий образ жизни, но при этом задний конец их раковины всегда возвышается над грунтом и омывается водой, необходимой животному для дыхания. Зарываясь в песок или ил, моллюски выбирают оттуда мелких беспозвоночных (фораминиферы, остракоды и др.), которыми питаются.
Тело билатерально симметрично (рис. 479). Складки мантии срастаются по брюшной стороне, образуя мантийную полость, которая имеет вид открытой с обоих концов трубки. В нее открываются задняя кишка и отверстия почек. Циркуляция воды в мантийной полости обеспечивается работой ресничного эпителия мантии. Вода втягивается через заднее отверстие и, отдав кислород тканям, через то же отверстие выводится наружу. Вместе с водой выносятся непереваренные остатки пищи, экскреты, а в период размножения и половые продукты. Специальных органов дыхания у лопатоногих нет.
Голова представляет вырост, на конце которого открывается рот и располагаются многочисленные нитевидные, утолщенные на концах усики, которые служат для осязания и захвата добычи. На расширениях усиков сосредоточены железистые клетки, выделяющие слизь, к которой, по-видимому, приклеиваются фораминиферы и другие мелкие животные, служащие лопатоногим пищей.
Нога приспособлена к копанию в грунте. Она заканчивается тупым конусом, у основания которого сидит пара боковых лопастей; их присутствие и дало основание для названия класса — лопатоногие. При передвижении в грунте нога сильно вытягивается вперед, боковые лопасти складываются, нога легко проникает в грунт. Затем боковые лопасти расправляются и, как якорь, закрепляют конец ноги, которая вслед за этим сильно укорачивается, подтягивая животное вперед.
Нервная система разбросанно-узлового типа. Органы чувств слабо развиты. Глаза редуцированы в связи с роющим образом жизни, статоцисты имеются. Пищеварительная система представлена глоткой с челюстями и радулой. Имеется парная печень. Кровеносная система лакунарная, сильно редуцирована; сердце лишено предсердий. Почки
с перикардием не сообщаются. Непарная половая железа открывается в правую почку. Лопатоногие раздельнополы. Яйца развиваются в морской воде. Дробление происходит по спиральному типу. Из яйца выходит типичная трохофорная личинка.
Лопатоногие известны с ордовика, хотя их ископаемые остатки встречаются редко. Число вымерших видов не превышает 200. Современные Scaphopoda распадаются на 2 семейства, различающиеся размерами и строением ноги. Преимущественно это южные, нередко тропические виды. В наших северных морях обычны довольно крупные Denta-lium entale и Siphodentalium lobatum, раковина которых достигает лишь нескольких миллиметров в длину.
КЛАСС V. ГОЛОВОНОГИЕ (CEPHALOPODA)

Рис. 480. Различные головоногие (по Хуну). А—Amphitretus pela-gicus (глубоководный плавающий осьминог); Б — Benthoctopus pro-fundorum (осьминог); В — Doratopsis sagitta (планктонный кальмар); Г — Lycoteuthis diadema (глубоководный пелагический кальмар со светящимися органами); Д — Cranchia scarba (планктонный кальмарчик); Е — Cirrothauma murrayi (донный, живущий на мягком грунте осьминог); Ж — Loligo edulis (пелагический кальмар)
|
| 8 |
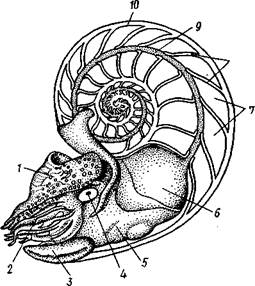
| Рис. 481. Кораблик Nautilus pompilius (подкл. Tetrabranchia) с распиленной раковиной (по Оуэну): 1 — головной капюшон, 2 — щупальца, 3 — воронка. 4 — глаз, 5 — мантия, 6 — внутренностный мешок, 7 — камеры, 8 — перегородки между камерами, 9 — сифон, 10 — стенка раковины |
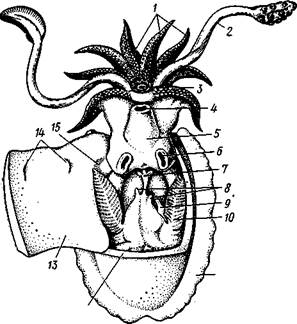
Класс головоногих (рис. 480) содержит около 700 живущих в морях видов наиболее сложно организованных, крупных, а иногда даже очень крупных (до 18м) моллюсков. Это свободноплавающие, реже ползающие, очень подвижные хищники, преимущественно обитающие в теплых морях. Тело их билатерально симметрично, с резким разделением на голову и туловище. Нога превращена в щупальца, или руки, которые вторично сместились на голову и окружают ротовое отверстие. Другая часть ноги представлена так называемой воронкой, лежащей на брюшной стороне у входа в мантийную полость. Раковина у примитивных форм наружная, многокамерная, у высших представителей — внутренняя, часто более или менее редуцированная. Нервная система и органы чувств достигают большого совершенства. Строение и физиология.Внешняя морфология. Голова несет на переднем конце рот, окруженный щупальцами. У немногих древних форм, например, кораблика Nautilus (рис. 481), имеется значительное и не вполне определенное количество тонких, червеобразных щупалец, которые служат\ для захватывания добычи и могут втягиваться в особые влагалища. Сильно разросшиеся влагалища дорзальных щупалец составляют так называемый капюшон — мускулистое образование, с помощью которого животное может закрывать вход в раковину. У высших головоногих всего 8 (отр. Octopoda) или 10 щупалец (отр. Dec.apoda), устроенных иначе, чем у наутилуса. Восемь щупалец Octopoda представлены широкими у основания, заостряющимися к концу мускулистыми выростами с продольным желобом на обращенной ко рту внутренней стороне. Эта сторона щупалец усажена многочисленными крупными дисковидны-ми присосками, позволяющими животному необычайно прочно присасываться ко дну и схватывать добычу.
У Decapoda кроме этих 8 щупалец есть еще 2 гораздо более длинных ловчих щупальца, расширенных на конце.
По бокам головы расположена пара очень крупных глаз. Туловище со всех сторон одето мантией. На спине она образует покровы самого туловища; на брюшной стороне она отделена от туловища мантийной полостью. На месте перехода туловища в голову мантийная полость сообщается щелевидным брюшным отверстием с внешней средой.
Для замыкания брюшной щели у высших головоногих есть особое приспособление в виде пары полулунных ямок на брюшной стороне туловища, соответственно которым на внутренней поверхности мантии ле-
жат два твердых, укрепленных хрящом бугра, или запонки. Действием мантийных мышц мантия прижимается к туловищу, запонки входят в полулунные ямки и как бы пристегивают мантию к телу, закрывая этим брюшную щель. Между обеими запонковыми ямками на брюшной стороне туловища помещается воронка, мускулистая коническая трубка, приросшая к туловищу. Расширенный задний конец воронки открывается в мантийную полость, суженный передний смотрит наружу. Воронка служит для движения. Когда мантийная щель закрыта, мантия прижимается к туловищу действием многочисленных находящихся в ней мышц и вода из мантийной полости с силой выталкивается наружу через переднее отверстие воронки, сообщая животному обратный толчок. Затем щель раскрывается и полость мантии заполняется окружающей водой. Далее происходит новое замыкание щели, сжимание мантии и новый толчок. Ритмические сокращения мантии и выталкивание воды служат, с одной стороны, в качестве дыхательного акта для смены воды в мантийной полости, с другой — для плавательного движения, которое совершается у головоногих толчками задним концом вперед. По бокам тела нередко образуются кожные складки, поддерживающие тело в равновесии и служащие плавниками.
В мантийной полости на брюшной стороне тела открывается порошица, по бокам ее расположены половые и почечные отверстия и ктенидии.
Нога, на первый взгляд, отсутствует, но это отсутствие лишь кажущееся. Нога Cephalopoda сильно видоизменена, и ей соответствует сово-
|
| 12 |
Рис. 482. Каракатица Sepia officinalis со вскрытой мантийной полостью, вид с брюшной стороны (по
Пфуртшеллеру):
Руки с присосками, 2 — ловчая рука, 3 — рот, 4 — отверстие воронки, 5 — воронка, 6 — хрящевые ямки запонок, 7 — анальный сосочек с анальным отверстием, 8 — почечные сосочки, 9 — непарный половой сосочек, 10 — жабры, //— плавник, 12 — линия отреза мантии, 13 — отогнутая мантия, 14 — хрящевые бугорки запонок. /5 — мантийный звездчатый ганглий
купность двух образований: воронки и щупалец. Гомология воронки ноге доказывается ее положением на брюшной стороне туловища, а также тем, что у древнего и примитивного Nautilus она не в виде трубки, а в виде желоба, раскрытого по срединной брюшной линии, т. е. очень напоминает ногу некоторых плавающих Gastropoda. У зародышей всех Cephalopoda воронка двухлопастная, и лопасти ее лишь вторично срастаются своими боковыми краями в трубку. Щупальца во время развития закладываются позади рта, на брюшной стороне тела, и лишь позднее надвигаются с двух сторон на голову, образуя околоротовой венец. Следовательно, по первичному положению щупальца соответствуют передней части ноги. Самым же важным доказательством указанного происхождения воронки и щупалец является иннервация их от педальных ганглиев.
|
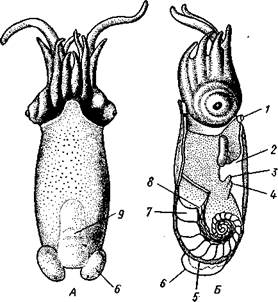
Раковина большей частью рудиментарна и хорошо развита лишь у наиболее древней из сохранившихся до сих пор форм, у кораблика Nauilus (см. рис. 481). Известковая наружная раковина наутилуса завита в одной плоскости на спинную сторону. Раковина обращена завитком вперед и замечательна тем, что полость ее разделена поперечными пере-
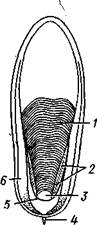
Рис.483. Двужаберный толстоногий моллюск Spirilla со спиральной многокамерной раковиной (из Натали). А — вид со спинной стороны; Б — продольный
разрез:
/ — воронка, 2 — мантийная полость. 3 — анальное отверстие, 4 — выделительное отверстие, S — орган свечения, 6 — плавник, 7 — раковина, 8 — сифон, 9 — участок раковины, покрытый
мантией
Рис. 484. Раковина каракатицы Sepia officinalis с брюшной стороны (по Мейеру)
ni_. tut. галииппа лазала inij,Di иь/укл I// / i-^'n+м j ^ vypiuuinv-ri v.ivpwnui \iiiy I'ltncyy j .
— сближенные перегородки спинной стороны фрагмокона. 2—боковой край проостракума,
— сифональная ямка, 4 — рострум, 5 — рудимент брюшной стенки сифональной трубки,
6 — задний кпяй ппоостпакума
6 — задний край проостракума
городками на ряд камер. Тело животного помещается лишь в последней, ближайшей к устью и самой большой камере (жилая камера), тогда как все прочие заполнены газом и некоторым количеством жидкости. При изменении объемных соотношений газа и жидкости животное может всплывать или погружаться на значительную глубину, используя раковину как своеобразный гидростатический аппарат. Посередине
каждой перегородки имеется по небольшому отверстию. Через них все камеры пронизываются тонким цилиндрическим выростом задней части туловища — сифоном. Самая первая и маленькая камера раковины называется эмбриональной. В палеозойских и мезозойских отложениях найдены остатки нескольких тысяч видов класса Cephalopoda с раковиной, построенной в общем по типу Nautilus. У современных головоногих раковина более или менее редуцируется, уменьшается в размерах и обрастает боковыми складками мантии, становясь внутренней. У интересного глубоководного рода Spirula (рис. 483) есть такая же, как у наутилуса, многокамерная раковина, но она так мала, что последняя камера ее содержит лишь небольшую часть животного и тело спирулы обрастает раковину со всех сторон.
У других представителей класса Cephalopoda, еще сохранивших раковину, она в виде известковой пластинки сложного строения, залегающей под покровами на спинной стороне туловища. Такова, например, раковина каракатицы Sepia (рис. 484). Спинная сторона такой раковины прикрыта тонким роговым листком (проостракум), а задний конец образует очень маленький конус, заканчивающийся известковым рожком (рострумом). Главную массу раковинной пластинки составляет множество плотных известковых пластиночек, отходящих от проостракума косо книзу (к брюшной стороне тела). Промежутки между пластиночками очень узки и заняты рыхлой известковой массой. Происхождение такой раковины выясняется при сравнении ее с раковиной вымерших мезозойских белемнитов Belemnites (рис. 485, В). Раковина их в виде длинного прямого конуса со множеством очень узких камер, разделенных перегородками. Такая раковина называется фрагмоконом. Перегородки пронизаны сифоном, проходящим ближе к брюшной стороне конуса. Его спинная сторона выдается вперед в виде тонкого листовидного рогового продолжения. Задний конец одет мощным коническим известковым фут-
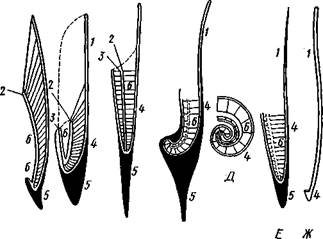
|
| J |
| В |
| Г |
Рис. 485. Схема строения раковины различных головоногих в сагиттальном разрезе; брюшная сторона раковины обращена налево. А — Sepia; Б — Belosepia; В — Belemnites (ископаемая) ; Г — Spirulirostra (ископаемая); Д — Spirula; Е — Ostra-coteuthis (ископаемая); Ж — Ommastrephes; 3 — Loligopsis (из Гешелера);
Дата добавления: 2015-01-24; просмотров: 2288;