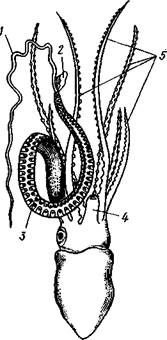
Раковина, просвечивающая сквозь покрывающие ее расширения руки (2), 3 — гектокотиль, 4 — глаз, 5 — воронка
nites был представлен громадным чертовым пальцем, а от конической раковины белемнита сохраняется, в основном, ее спинная часть, тогда как брюшная почти полностью исчезла. Перегородки спинной части фрагмокона чрезвычайно сблизились (косые пластиночки раковины Sepia) и промежутки между ними заполнились известью, так что эта часть раковины приняла вид слоистой пористой известковой пластинки. Задний конец фрагмокона сохранил) однако, не только спинную, но и брюшную стенку (рис. 485, А), образуя маленькую вороночку — сифональную ямку (см. рис. 484), полость которой представляет собой сохранившийся задний участок сифона.
У некоторых других современных головоногих, например у кальмара (Lotigo), от всей раковины сохраняется лишь спинной роговой листок (проостракум), скрытый под покровами, а у большинства Octopoda раковина совершенно исчезает. Своеобразной «раковиной» обладают Argonauta из Octopoda (рис. 486). Раковина есть только у самок. Она спирально закручена, но не разделена на камеры и очень тонка. Это вторичное образование, не имеющее ничего обшего с раковиной прочих моллюсков, и выделяющееся не мантией, а эпителием особых лопастей щупалец. Раковина Argonauta служит для вынашивания яиц.
Покровы. Кожа головоногих состоит из однослойного цилиндрического эпителия и подстилающего его слоя соединительной ткани. Cephalopoda обнаруживают замечательную по быстроте и резкости смену окраски, которая находится под контролем нервной системы. Смену цветов вызывает изменение формы многочисленных пигментных клеток, или хроматофоров, залегающих в соединительнотканом слое кожи.
Внутренний скелет. Интересно, что кроме наружного скелета или раковины у головоногих есть еще особый внутренний скелет, служащий
. 490
главным образом для защиты центральной нервной^системы. Он образован хрящом, сходным с таковым позвоночных. Хрящ в виде широкого кольца охватывает головное скопление ганглиев, образуя головную хрящевую капсулу, которая дает от себя выросты, окружающие глаза и ста-тоцисты. Кроме того, опорные хрящи развиты в запонках, в основании щупалец, внутри плавников. Как видно из описания, головная капсула в физиологическом отношении соответствует черепу позвоночных.
|
| 1 |
| 15 |
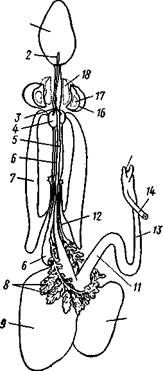
Пищеварительный канал у Cephalopoda устроен очень сложно (рис. 487). Рот лежит в центре венца щупалец и ведет в сильно развитую, мускулистую глотку. В глотке имеется язык с радулой, но терка играет в захвате и размельчении пищи второстепенную роль. Главное значение имеют две толстые роговые челюсти — спинная и брюшная, загнутые крючковидно и похожие на клюв попугая. В глотку впадают протоки одной или двух пар слюнных желез. Секрет их содержит ряд гидролитических ферментов, обеспечивающих расщепление, полисахаридов и белков. Выделения задней пары желез ядовиты. Длинный пищевод нередко образует расширение, зоб (отр. Octopoda). Пищевод переходит в мускулистый мешковидный желудок, занимающий заднюю часть пищеварительного комплекса. От той же передней стороны желудка, куда впадает пищевод, начинается тонкая кишка, которая направляется вперед. Она ведет к порошице, лежащей на особом сосочке на брюшной стороне туловища (внутри мантийной полости).
| 10 |
| Рис. 487. Пищеварительная система каракатицы Sepia officinalis, вид с брюшной стороны (по Резелеру и Лампрехту): / — глотка, 2 — общий слкшной проток, 3 — слюнной проток, 4 — задняя слюнная железа, 5 — пищевод, 6 — головная аорта, 7 — печень, 8 — поджелудочная железа, 9 — желудок, 10 — слепой мешок желудка, // — тонкая кишка, 12 — печеночный проток, 13 — прямая кишка, 14 — проток чернильного мешка, 15 — анус, 16 — головная хрящевая капсула (разрезана), 17 — полость капсулы статоциста, 18 — нервное кольцо (разрезано) |
В желудок впадают протоки двухлопастной, реже простой печени, секрет которой содержит широкий спектр пищеварительных ферментов. Печеночные протоки большей частью усажены многочисленными небольшими железистыми придатками, выделяющими ферменты, гидролизующие полисаха-риды. Эти придатки называются поджелудочной железой. Как видно, печень и поджелудочная железа головоногих отнюдь не соответствуют в точности одноименным образованиям у позвоночных. В заднюю кишку, перед самой порошицей, открывается проток так называемого чернильного мешка. Это большая грушевидная железа, выделяющая черную, как чернила, жидкость. Несколько капель этого секрета достаточно, чтобы замутить большое количество воды. Выбрасывая жидкость через порошицу, а далее через отверстие воронки наружу, Cephalopoda окружают себя в воде темным облаком и под покровом его ускользают от преследования. Все головоногие — хищники; они нападают на различных ракообразных, а иногда и на рыб, которых схватывают щупальцами и убивают челюстями и ядом слюнных желез.
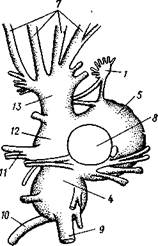
Нервная система головоногих достигает высокой сложности строения (рис. 488, Б). Ганглии ее велики и образуют общую окологлоточную нервную массу. Лишь на разрезах можно различить состав этой массы из нескольких ганглиев: ясно различимы парный церебральный ганглий и большой висцеральный. Что касается пары педальных ганглиев, то характерно подразделение каждого из них на два нервных узла: брахиаль-ный, или ганглий щупалец, и инфундибулярный, или ганглий воронки.
От заднего отдела ганглиозной массы отходят 2 крупных мантийных нерва, которые образуют на внутренней поверхности мантии, по бокам в передней части туловища, 2 больших звездчатых гзнглия. У более примитивных Tetrabranchia (Nautilus) центральная нервная система устроена значительно проще (рис. 488, А); она состоит из трех коротких нервных дуг, одной надглоточной и двух подглоточных, причем по бокам пищевода все дуги соединяются между собой. Дуги сплошь усеяны ганг-лиозными клетками и носят характер нервных тяжей (как у Amphineura).
Органами чувств головоногие снабжены богато. Для чувства обоняния служат либо осфрадии, расположенные у основания жабр (Nautilus), либо пара расположенных под глазами обонятельных ямочек (Dib-ranchia), небольших кожных углублений, выстланных чувствительным эпителием и иннервируемых от церебральных ганглиев. Острота обоняния довольно велика, это показывают опыты над ослепленными осьминогами (Octopus), которые чувствуют положенную в аквариум рыбу на .расстоянии 1,5 м.
Имеется пара статоцистов, заключенных внутри хрящевой головной капсулы. Удаление их ведет к утрате способности ориентироваться в пространстве.
Органы зрения Tetrabranchia могут быть сведены к типу строения крупных глазных ямок, полость которых маленьким отверстием еще со-
|

|
Рис. 488. Центральная нервная система головоногих. А — Tetrabranchia (Nautilus), вид спереди (из Кестнера, по Гриффину); Б — Dibranchia (Sepia), вид сбоку (из Кестнера,
по Хиллигу, с изменениями):
/— Оуккальный ганглий, 1 — педальный ганглий, 3 — оптический нерв, 4 — висцеральный ганглий, 5 — церебральный ганглий, 6 — нерв, идущий к капюшону и щупальцам, 7 — нервы рук, 8 — место отхождения оптического нерва, 9 — мантийный нерв, 10 — нерв, идущий к внутренностям, 11 — нерв воронки, 12 — инфундибулярный ганглий, 13 — брахиальный ганглий, (12 и 13 — производные педального ганглии)
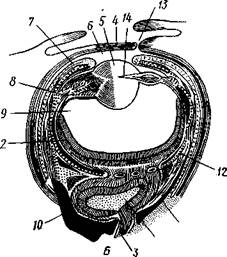
общается с внешней средой (рис. 489, Л). У всех прочих Cephalopoda глаза носят характер очень крупных, замкнутых, сложно устроенных глазных пузырей (рис. 489, 6). Строение их легче всего понять, прослеживая историю развития глаза. Сначала у зародыша образуется первичная глазная ямка (стадия Nautilus), которая отшнуровывается от кожи и дает глазной пузырь. Глубокая часть стенки пузыря превращается в
|

|
| 10 |
| 11 |
Рис. 489. Глаза головоногих. А — разрез глаза Nautilus (Tetrabranchia) (из Гешелера); Б — разрез глаза Sepia officinalis (Dibranchia) (по Генсену, с
изменениями):
Полость глазной ямки, сообщающейся с внешней средой, 2 — сетчатка, 3 — зрительный нерв, 4 — роговица, 5 — хрусталик, 6 — передняя камера глаза, 7 — радужина, 8 — ресничный мускул, при сокращении приближающий хрусталик к сетчатке, 9 — стекловидное тело, 10 — глазные отростки хрящевой капсулы, // — оптический ганглий, 12 — склера (хрящевая оболочка глаза), 13 — наружное отверстие камеры глаза, /4 — эпителиальное тело
сетчатку, тогда как наружная, прилегающая к коже часть дает в совокупности с наружным эпителием двухслойное эпителиальное тело. Вокруг глаза появляется кольцевая складка кожи — радужина. Она нарастает над глазом в виде свода, но не вполне смыкается, так что в центре свода остается отверстие — зрачок. Между тем обоими слоями эпителиального тела выделяется одним слоем кнаружи, другим — в полость глазного пузыря по прозрачному полушарию. Совокупность обоих полушарий дает шаровидный хрусталик, перерезанный тонкой двойной пластинкой эпителиального тела. Тем временем на глаз нарастает новая кольцевидная складка кожи — роговица. У большинства головоногих свод, образуемый роговицей над радужинным сводом, не вполне смыкается, сохраняя эксцентрическое отверстие. Сетчатка глаз состоит из очень длинных (иногда до 0,5 мм) клеток; к ней подходит мощный зрительный нерв, образующий под самым глазом очень крупный зрительный ганглий из нескольких ярусов нервных клеток.
Глаза головоногих обладают аккомодацией, которая совершается, однако, не изменением кривизны хрусталика (как у человека), а его приближением или удалением от сетчатки; для этого служит особый ресничный мускул, прикрепленный к экватору хрусталика. Кроме того, в глазу Cephalopoda имеются приспособления к видению при более сильном и более слабом освещении.
Острота зрения животного отчасти зависит от числа воспринимающих свет элементов. В глазу головоногих этих элементов очень много. Так, у каракатицы Sepia на 1 мм2 сетчатки приходится 105000 клеток; у кальмара Loligo — даже 165000. Все это убеждает нас в большой сложности строения глаз головоногих, которые в этом отношении не уступают даже глазам позвоночных.
У многих головоногих (особенно у глубоководных) в коже имеются особые органы свечения (см. рис. 480,Г), по строению несколько похожие на глаза. Так, у Lycoteuthis diadema 22 подобных органа, из которых часть светится ультрамариновым, часть — светло-голубым, а некоторые— рубиново-красным фосфорическим светом.
Органы дыхания. Головоногие имеют настоящие ктенидии. У всех Dibranchia их 2, у Tetrabranchia (Nautilus) —4. Ктенидии расположены в мантийной полости симметрично по бокам туловища (см. рис. 491, 482). Жабры двоякоперистые, свободными концами направлены вперед.
Обмен воды в мантийной полости обеспечивается сокращением мантийных мышц и деятельностью воронки. По количеству жабр Cephalopoda делятся на два подкласса: четырехжаберные (Tetrabranchia) и дву-жаберные (Dibranchia).
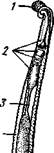
Кровеносная система. Сердце всех головоногих состоит из одного желудочка, в который открываются с боков два (подкл. Dibranchia; рис. 490) или четыре (Tetrabranchia) предсердия. От переднего и заднего концов желудочка отходят две аорты: головная и внутренностная.
Головная аорта идет рядом с пищеводом и дает ветви к голове и щупальцам. Внутренностная снабжает кровью кишечник и половые органы. Артерии разбиваются на сеть капилляров, из которых берут начало вены.- Вены рук собираются в голове в одну кольцевую вену, от кото-

|
| 12 |
Рис. 490. Центральная часть кровеносной системы, жабры и почки каракатицы Sepia (из Кестнера):
/ — головная вена, 2 — наружное отверстие почек. 3—полая вена, 4—почка, 5 — уносящий жаберный сосуд, 6 — приносящий жаберный сосуд, 7 — венозное (жаберное) сердце, 8—перикардиальная железа, 9 — перикардиальная полость, 10 — внутренностная аорта, ;/ —желудочек, 12—предсердие, 13 — реио-перикардиальное отверстие, 14 — головная аорта (черным обозначены сосуды, по которым циркулирует венозная кровь, белым — сердце и сосуды с артериальной кровью)
|
| 5--М |
|
рой рядом с головной аортой, но назад, идет крупная головная вена. Головная вена (рис. 490) делится на заднем конце на 2 или 4 (смотря по числу жабр) приносящих жаберных сосуда (полые вены), которые принимают еще кровь из мантии и несут венозную кровь к жабрам. Перед вступлением в жабры они образуют сократимые мускулистые расширения, так называемые венозные (жаберные) сердца, которые ритмическими сокращениями способствуют поступлению крови в жабры. Кровь окисляется в капиллярах жабр и затем по уносящим сосудам изливается в предсердия сердца. Капилляры вен и артерий в коже и мышцах непосредственно переходят друг в друга, лишь в некоторых местах между ними остаются лаку-нарные пространства, что делает кровеносную
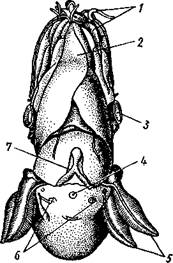
Рис. 491. Мантийный комплекс Nautilus. Вид животного, вынутого из раковины, с брюшной стороны, мантия удалена (по Тиле):
/ — щупальца, 2 — воронка, 3 — глаз, 4—> анальное отверстие, 5 — жабры, 6 — экскреторные отверстия, 7 — пенис
Рис. 492. Сперматофор
каракатицы Sepia (по
Мильн-Эдвардсу):
/ — закрученный конец спер-матофора, 2 — различные части семяизвергательного аппарата, 3 — соединительный тяж, 4—хитиноидная оболочка, 5 — резервуар с семенем
систему головоногих почти замкнутой. Кровь содержит гемоцианин — богатое медью соединение, физиологически соответствующее гемоглобину крови позвоночных; на воздухе синеет.
Выделительная система состоит из 4 (подкл. Tetrabranchia; рис. 491) или 2 (подкл. Dibranchia; см. рис. 490) почек. Наружные отверстия их лежат по бокам порошицы, на особых сосочках (см. рис. 482), внутренние концы почек открываются, как всегда, в перикардиальный отдел це-лома. Почки представляют обширные мешки, иногда (отр. Decapoda) срастающиеся друг с другом по срединной линии тела.
Проходящие в тесном соседстве с почками приносящие жаберные сосуды (с венозной кровью) образуют многочисленные бахромчатые
слепые выпячивания (венозные придатки), которыми вдаются в стенки почек, этим облегчается извлечение почками продуктов обмена веществ из крови.
Половая система. Головоногие моллюски раздельнополы, иногда с резким половым диморфизмом. Так, у Argonauta самец гораздо меньше самки (см. рис. 486).
Половая железа непарна и залегает в задней части туловища, в половом участке целома. Половые клетки скапливаются в целоме и выводятся через половые протоки. Протоки первично парны, хотя у многих головоногих остается только левый проток. Парность протоков сохраняется у самцов и у самок Nautilus, у Octopoda и части Decapoda.
Протоки обладают сложным и варьирующим строением. У самца ка« ракатицы Sepia имеется семяпровод, расширяющийся в семенной пузырек, железистый эпителий которого служит для образования спермато-форов. За пузырьком семяпровод снова суживается и переходит затем в широкую сперматофорную сумку, которая и открывается сбоку от порошицы половым отверстием. Живчики головоногих склеиваются в окруженные плотной оболочкой пакеты, или сперматофоры (рис. 492). Спер-матофоры наполнены живчиками и имеют особый канал для их выхода, заткнутый сложно устроенной пробочкой.
Женские протоки состоят из короткого яйцевода с впадающей в него небольшой яйцеводной железой. Кроме того, . независимо от полового
|
отверстия, но по соседству с ним в мантийную полость открываются выводные каналы двух парных и одной непарной нидаменталь-ных желез, секрет которых служит для образования яйцевых оболочек.
| Рис. 493. Самец осьминога Ocythoe catenulata с гектокоти- лизированной рукой (по Пель- знеру): I — концевая нить гектокотнля, 2 — мешочек гектокотиля, внутри которого развивается нить. 3 — гектоко-тилизированная рука, 4 — воронка, 5 — руки |
Оплодотворение чаще всего происходит в мантийной полости самки. Роль копуля-тивного органа играет одно из щупалец, у самцов оно более или менее сильно отличается от остальных иным развитием присосок и по присутствию на нем специального ложкообразного придатка (рис. 493). При помощи этого щупальца самец вводит сперматофоры в мантийную полость самки. Особенно замечательным устройством обладает половое щупальце у самца Argonauta (см. рис. 486) и некоторых других близких к нему родов. Щупальце сначала образуется внутри особого кожистого мешка, в котором оно свернуто в спираль. Конец полового щупальца вытянут в длинную нить. Щупальце содержит полость, сообщающуюся с внешней средой двумя отверстиями: одним у основания, другим на конце. Когда щупальце вполне сформируется, то облекающий его мешок лопается и оно расправляется. Полость щупальца (еще не выяснено, каким образом) наполняется сперматофорами. При спаривании половое щупальце Argonauta отрывается, надолго сохраняет способность к движению и проникает целиком в мантийную полость самки, где оно вводит сперматофоры в женское половое отверстие.
Когда оторвавшиеся щупальца самцов были впервые найдены в манти-й-ной полости самок, они были приняты за паразитов, и Кювье дал им родовое название Hectocotilus; это название и до сих пор сохранилось за половым щупальцем головоногих, которое называют «гектокотилизиро-ванным».
Развитие.Эмбриональное развитие Cephalopoda отличается многими особенностями, определяемыми главным образом богатством яиц желтком. Он занимает почти все яйцо, его нет только на анимальном полюсе. Дробление вследствие обилия желтка частичное, причем делится на бластомеры лишь анимальный полюс; здесь образуется округлая пластинка, или диск, зародышевых клеток (частичное дробление головоногих называется дискоидальным). Развитие зародыша идет главным образом за счет анимального диска, на котором зародыш расположен так, что головным концом он обращен к желтку, а задним торчит свободно. Желток обрастается эктодермой и мезодермой зародыша, но остается довольно резко обособленным от последнего в виде желточного мешка. По мере роста зародыша желток потребляется, и желточный мешок уменьшается в размерах. Щупальца закладываются позади рта в виде двух боковых рядов, воронка имеет вид двухлопастного, открытого на брюшной стороне образования и лишь позднее срастается в трубку. Все развитие протекает внутри оболочки яйца.
Палеонтология.Наиболее древними считаются известные с кембрия Nautiloidaea — родичи современного наутилуса. У первых наутилоидей раковина еще не была спирально закрученной и имела вид конической трубки, поделенной на камеры и пронизанной узким сифоном (Orthoce-ras). Такие или слегка согнутые наподобие рога раковины (Cyrtoceras) были особенно многочисленны в ордовике. Позднее появились вытеснившие их виды с раковиной, скрученной спирально, как у современного наутилуса, сохранившегося с триаса до наших дней.
В ордовике были распространены гигантские Endoceras (представители особого отряда), прямая раковина которых, тоже поделенная на камеры, достигала 4—5 м длины. В широком сифоне, занимавшем около '/з диаметра раковины этих моллюсков, помещалась значительная часть органов животного. Эндоцерасы вели придонный образ жизни.
|
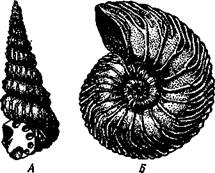
| Рис. 494. Аммониты. А — Turrilites catenatus, редкая среди аммонитов форма закручивания раковины в коническую спираль (по Куку); Б — планоспиральная раковина Cardiacs-ras cordatum — хорошо сохранилась скульптура стенки раковины (по Бо« дылеаскому) |
В девоне появляется небольшая группа относительно мелких головоногих с прямой или слегка согнутой раковиной — бактриты. По современным представлениям эти животные, просуществовавшие до конца палеозоя, дали начало двум большим ветвям развития головоногих — аммонитам, известным с девона, и белемнитам. Это чрезвычайно многочисленная и богатая пред-ствителями группа отрядов, существовавшая в палеозое и мезозое. В кайнозое аммониты отсутствуют, они вымерли в конце мелового периода.
Аммониты обладали спирально закрученной многокамерной раковиной, обороты которой в большинстве случаев располагались в одной плоскости. Раковины аммонитов от-
личались разнообразием формы (рис. 494), наружной скульптуры и размеров. Наряду с очень мелкими видами встречались настоящие гиганты, например Pachydiscus, планоспиральная раковина которого в диаметре достигала 2 м. Сведений о внутреннем строении аммонитов почти нет. По-видимому, они обладали вытянутым червеобразным телом. Полагают, что у них было 10 щупалец, чернильный мешок и, возможно, всего 2 ктенидия. Относятся ли аммониты к четырехжаберным или к двухжаберным, до сих пор еще не установлено. Вероятно, аммониты были хищниками и вели плавающий образ жизни. Они населяли моря земного шара на протяжении сотен миллионов лет. Бесчисленные остатки этих головоногих широко используются для определения геологического возраста слоев земной коры.
Все рассмотренные группы головоногих обладали хорошо развитой раковиной, в которую целиком могло втягиваться тело животного. На этом основании палеонтологи объединяют их под названием наружнора-ковинных. Начиная с карбона, появляются головоногие, раковина которых снаружи обрастает мантией. Это внутреннераковинные, которые, как и аммониты, ведут начало от бактритов, но являются уже, несомненно, двужаберными. Эта ветвь головоногих достигла высшего расцвета в кайнозое. Сюда относятся современные кальмары и осьминоги. Среди ископаемых представителей этой группы особенно своеобразными были белемниты. Они появились в карбоне, достигли расцвета в юре и мелу и полностью исчезли к середине палеогена. Это были, по-видимому, плавающие моллюски. Задний конец их конической раковины (с. 489) имел вид массивного известкового конуса, иногда достигавшего значительной длины. Именно эта часть их раковины — рострум, наиболее часто встречается в мезозойских отложениях.
Ископаемые головоногие по количеству видов несравненно многочисленнее, чем современные. Их насчитывают до 11 000 видов.
Экология. Встречаясь исключительно в морях, головоногие ведут различный образ жизни. Большинство их относится к пелагическим животным. Такие виды, как Loligo, имеют торпедообразное, заостренное на заднем конце (который при движении направлен вперед) тело с хорошо развитыми плавниками.
У некоторых донных осьминогов (Cirrothauma) между щупальцами развивается тонкая перепонка, так что животное принимает вид диска, лежащего на дне. Другие головоногие живут на дне, нередко прячась в расщелины между камнями. Глубоководные головоногие обладают иногда торчащими, наподобие телескопов, глазами.
Головоногие — нередко весьма крупные животные. Самые мелкие из них измеряются несколькими сантиметрами, но на больших глубинах водятся настоящие гиганты.
У подобных форм такая мускульная сила, что они очень редко попадаются в руки человеку, да и то лишь по счастливой случайности. Так, однажды удалось выловить извергнутые умирающим кашалотом во время агонии остатки съеденного им громадного головоногого Architeuthis; сохранившееся щупальце этого животного имеет длину около 10 м, следовательно, общая длина Architeuthis с вытянутыми ловчими щупальцами достигала 18 м.
Практическое использование головоногих. Многие Cephalopoda служат предметом промысла. Каракатицы, кальмары и осьминоги употребляются в пищу в свежем, сушеном и консервированном виде. Их мясо по калорийности и вкусовым качествам не уступает говядине. Используется и секрет чернильного мешка каракатиц и кальмаров. После высуши-
вания и обработки едким калием осадок секрета дает акварельную краску сепию. Из этого же осадка приготовляют и натуральную китайскую тушь. Во всех странах мира ежегодно вылавливается около миллиона тонн головоногих моллюсков.
Классификация.Современные головоногие делятся на два подкласса.
ПОДКЛАСС 1. ЧЕТЫРЕХЖАБЕРНЫЕ (TETRABRANCHIA)
Сюда принадлежат наиболее древние и примитивные формы, обладающие четырьмя жабрами, четырьмя предсердиями, четырьмя почками и значительным количеством щупалец; воронка в виде открытого желоба; раковина наружная, многокамерная. Че-тырехжаберные представлены несколькими группами отрядов (надотрядами), среди которых мы назовем лишь главнейшие.
Надотряд 1. Наутилоидеи (Nautiloidea).Вымирающая группа, к которой в настоящее время принадлежит один род Nautilus — кораблик (см. рис. 481) с небольшим количеством видов, обитающих в Индийском и Тихом океанах. Ископаемые формы: Orthoceras.
Надотряд 2. Аммониты (Ammonoidea).Группа, целиком вымершая в меловом периоде. Представители: Turrilltes, Ammonites (рис. 494). Внутреннее строение аммонитоп плохо известно, поэтому не ясно их систематическое положение. Устройство раковины приближает их к четырехжаберным.
ПОДКЛАСС II. ДВУЖАБЕРНЫЕ (D1BRANCHIA)
Головоногие, обладающие двумя жабрами, двумя предсердиями и двумя почками; воронка в виде трубки, раковина внутренняя, более или менее редуцированная.
Отряд 1. Десятиногие (Decapoda).Имеют 10 щупалец, из них одна пара длинных, ловчих. Хорошие пловцы с вытянутым торпедообразным телом с боковыми плавниками. Представители: обыкновенная каракатица Sepia officinalis, гигантский кальмар Archi-teuthis, кальмар Loligo и др. (см. рис. 480). Ископаемые формы: Belosepia, Spirulirostra,
К десятиногим относится вымершая в эоцене группа белемнитов (Belemnoidea) с прямой конической раковиной и массивным рострумом. Представитель: Belemnites. Некоторые авторы выделяют белемнитов в самостоятельный отряд Belemnitida.
Отряд 2. Восьминогие (Octopoda).Большей частью донные формы с мешковидным телом и вполне редуцированной раковиной; щупалец 8, ловчих щупалец нет. Представители: аргонавт Argonauta, осьминоги Octopus и др. (см. рис. 480).
Дата добавления: 2015-01-24; просмотров: 1611;