Ядро, 2 — стрекательная капсула, 3 — книдоциль, 4 — стрекательная нить с шипиками, 5 — шипы, 6 — жгутик, 7 — микроворсинки
горий клеток (см. рис. 91, В). Основу ее составляют эпителиально-мускульные пищеварительные клетки, кроме них имеются особые железистые клетки. Мускульные отростки энтодермальных эпителиально-мускульных клеток расположены поперечно по отношению к продольной оси тела. При сокращении их тело гидры суживается, становится тоньше, т. е. они антагонисты эктодермальных эпителиально-мускульных клеток.
Эпителиальная часть энтодермальных клеток, направленная в сторону пищеварительной полости, несет 1—3 жгута и способна образовывать псевдоподии, которыми захватываются мелкие пищевые частицы. Таким образом, у кишечнополостных, как и у губок, имеет место внутриклеточное переваривание пищи — признак примитивной организации. Однако наряду с этим железистые клетки энтодермы выделяют пищеварительные соки непосредственно в гастральную полость, где также происходят процессы пищеварения, т. е. у гидры сочетаются внутриклеточное и полостное пищеварение. Гидра питается различными мелкими животными, преимущественно рачками (дафнии, циклопы).
Мезоглея у гидры и других гидроидных полипов в виде тонкой бесструктурной пластинки — базальной мембраны, залегающей между эктодермой и энтодермой.
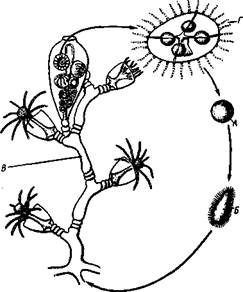
Размножение и развитие.Гидры размножаются бесполым и половым путями. Бесполое размножение состоит в почковании. Приблизительно на уровне середины тела гидры имеется так называемый пояс почкования. Здесь время от времени образуется бугор (почка), вырастающий и образующий на вершине новый рот и зачатки щупалец. Почка у основания перешнуровывается, падает на дно и переходит к самостоятельной жизни. Иногда почкование идет так энергично, что еще до отрывания первой почки на гидре успевают образоваться две-три другие (рис. 94).
Половым способом гидра начинает размножаться с приближением холодов. Гидры раздельнополы, но встречаются и гермафродитные виды, причем половые клетки их происходят из эктодермы, что характерно для всех Hydrozoa. Некоторые интерстициальные клетки эктодермы или непосредственно превращаются в яйца, или же многократным делением
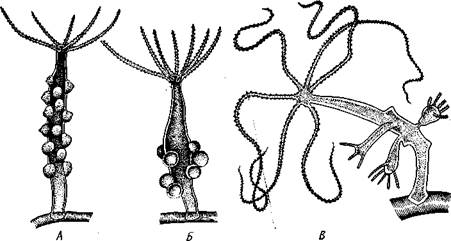
Рис. 94. Hydra oligactis при небольшом увеличении. А — с мужскими гонадами; Б — с женскими гэнадами; В — почкующаяся гидра (по Полянскому)
дают скопления живчиков (сперматозоидов). В этих местах на теле гидры эктодерма вздувается в виде бугорков. Яйца располагаются ближе к основанию гидры, а бугорки со сперматозоидами (мужские гонады) — к ротовому полюсу. Яйцо оплодотворяется в теле матери еще осенью и окружается плотной оболочкой, потом гидра погибает, а яйца остаются в покоящемся состоянии до весны, когда из них развиваются новые гидры.
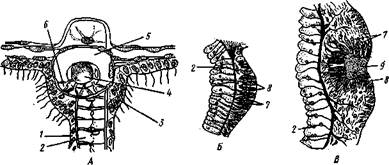
Морские гидроидные полипы.Лишь очень немногие гидроидные полипы наподобие гидры ведут одиночный образ жизни. Большинство образует колонии, состоящие из множества особей (рис. 95). Формирование колоний становится легко понятным из сравнения с гидрой. Представим себе, что почки, образовавшиеся на теле гидры, не отрываются от нее, а остаются с ней в постоянной связи и сами начинают почковаться, не отделяя от себя образующихся дочерних полипов. Получается комплекс особей, сидящих как бы на общем стволе и его побочных ветвях.
Колония чаще всего имеет вид деревца или кустика. Основание общего ствола колонии обыкновенно дает стелющиеся по субстрату отростки, похожие на корни растений и служащие для прикрепления колонии. Ствол ветвится, на ветвях сидят отдельные особи колонии — гидранты;
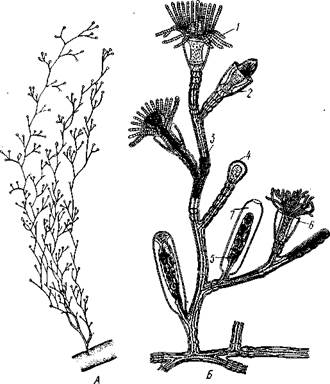
Рис. 95. Гидроид Obelia. A — колония (слегка увеличено); Б — отдельная веточка колонии (несколько схематизировано, часть особей колонии изображена в разрезе) (по Абрикосову):
1 — гидрант в расправленном состоянии, 2 — сократившийся гид
рант, 3 — тека, 4 — почка, 5 — бластостиль с развивающимися
медузами, 6 — гидротека, 7 — гонотека (участок теки, одевающий
ф бластостиль}
|
каждая особь соответствует как бы одной почке гидры и напоминает гидру. Гистология гидранта в общем отвечает таковой гидры, но гаст-ральная полость гидрантов продолжается в полость, или капал, проходящий через весь ствол и ветви колонии. Таким образом, гастральные полости всех гидрантов сообщаются между собой, так что пища, захваченная отдельными гидрантами, может затем в переработанном виде распределяться по всей колонии. Эктодермальный эпителий ствола выделяет на поверхности особую
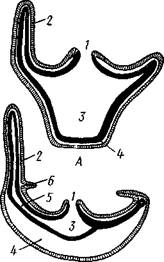
Рис. 96. Сравнение строения гидроидного полипа (Л) и гидроидной медузы (Б), перевернутой ротовым отверстием кверху (по Холодков-
скому):
I — рот, 2 — щупальца, 3 — гас-тральная полость, 4—мезоглея, 5 — радиальный канал, 6 — парус
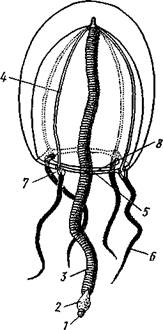
Рис. 97. Гидроидная медуза Sarsia (рис. А. Наумова):
J — рот, 2 — ротовой стебелек с расположенной на нем гонадой (3), 4 — радиальные каналы, 5 — кольцевой канал, 5 — щупальца, 7 — глазки, 8 — парус
органическую оболочку — теку, которая придает ему большую устойчивость. Тека доходит до основания гидрантов (подотряд Athecata), а иногда продолжается и на них самих (подотряд Thecaphora) в виде защитного колпачка, или гидротеки.
Размножение морских гидроидных полипов, образование и строение медуз.Сами полипы способны лишь к бесполому размножению почкованием, в гидрантах половые железы не образуются. Они формируются лишь в специальных, возникающих на колонии тоже путем почкования половых особях, медузах, переходящих к свободноплавающему образу жизни.
Сначала на известных местах стебля колонии появляется вырост, напоминающий зачаток гидранта. Этот вырост сильно вытягивается и превращается в полый столбик, видоизмененный полип — бластостиль, по бокам которого выпочковываются зачатки медув. Каждый зачаток постепенно превращается в молодую медузу, которая отрывается от бла-стостиля и уплывает. Медузка растет, развивает в себе половые клетки и
приступает к половому размножению. Иногда медузы выпочковываются поодиночке, без бластостиля.
Медузы, за исключением некоторых отклонений в организации пищеварительной системы, построены по той же схеме, что и полипы, но часто сильно сплющены в плоскости, перпендикулярной к главной оси тела (рис. 96).
Медуза имеет вид колокола или зонтина; наружная выпуклая сторона называется эксумбреллой, внутренняя вогнутая — субумбреллой (рис. 97). Посредине последней выдается более или менее длинный ротовой стебелек со ртом на свободном конце. Рот ведет в пищеварительную, или гастральную, полость, состоящую из центрального желудка и расходящихся от него к краям зонтика радиальных каналов в числе, равном четырем или кратном четырем, и соединенных в толще мезоглеи сплошной энтодермальной пластинкой. На краю зонтика все радиальные каналы сообщаются между собой посредством кольцевого канала. Желудок и каналы в совокупности образуют гастроваскулярную (т. е. кишечносо-судистую) систему. По свободному краю зонтика прикреплена тонкая кольцевидная мускулистая перепонка, суживающая вход в полость колокола. Она называется парусом и является характерной особенностью гидроидных медуз, отличающей их от медуз, принадлежащих к Scypho-zoa. Парус играет важную роль при движении медуз. На краю зонтика расположены, кроме того, щупальца. Они, подобно радиальным каналам, имеются в определенном числе, чаще всего кратном четырем.
Вследствие правильного расположения радиальных каналов и щупалец лучистая симметрия медуз ярко выражена.
Тело медуз характеризуется сильным развитием мезоглеи, которая очень утолщается и содержит большое количество воды, приобретая студенистый желеобразный вид. Благодаря этому все тело медуз почти стекловидно и прозрачно. Прозрачность, свойственная очень многим планктонным животным, рассматривается как особый род покровительственной окраски, укрывающей животное от врагов.
|
| Рис. 98. Органы чувств гидромедуз. А — основание щупальца и орган равновесия медузы Obelia (по О. и Р. Гертвигам); Б — глазное пятно медузы (из Хайман); В — глаз медузы (по Линко): / — эктодерма щупальца, 2—энтодерма, 3 — основание щупальца, 4 — стато-лит, 5 — полость статоциста, 6 — чувствительные клетки, 7 — ретинальиые клетки, S — пигментные клетки, 9 — хрусталик |
Нервная система медуз устроена значительно сложнее, чем у полипов. У медуз кроме общего подкожного нервного сплетения по краю зонтика наблюдаются скопления ганглиозных клеток, которые вместе с отростками образуют сплошное нервное кольцо. От него иннервируются мышечные волокна паруса, а также особые органы чувств, расположенные по краю зонтика. У одних гидроидных медуз эти органы имеют вид
глазков, у других — так называемых статоцистов, или органов равновесия (рис. 97, 98).
Глаза медуз в наиболее примитивном виде устроены по типу простых глазных пятен. У основания некоторых щупалец имеется небольшой участок эктодермального эпителия, состоящий из клеток двух родов. Одни из них высокие — чувствительные, или ретипальные, клетки; другие содержат многочисленные бурые или черные зерна пигмента и чередуются с чувствительными клетками, согюкупность которых отвечает сетчатке глаза высших животных. Присутствие пигмента вообще характерно для органов зрения во всем животном царстве.
Несколько более сложно устроены глазные ямки, где пигментированный участок эпителия лежит на дне небольшого ппячивания покрова. Подобный уход глаза с поверхности тела вглубь предохраняет его от различных чисто механических раздражений, например трения о воду, прикосновения посторонних предметов и т. п. Кроме того, впячивание глаза ведет к увеличению поверхности светочувствительного слоя и количества ретинальных клеток. Наконец, у некоторых медуз полость глазной ямки заполняется прозрачным выделением эктодермы, получающим вид лучепрсломляющей чечевицы (линзы). Таким путем возникает хрусталик, концентрирующий световые лучи на сетчатке глаза.
Органы равновесия могут быть устроены различно: в виде чувствительных щупалец, но чаще всего в виде глубоких эпителиальных ямок, которые могут отшнуровываться от поверхности тела и превращаться в замкнутые пузырьки, или статоцисты (рис. 98). Пузырек выстлан чувствительным эктодермальным эпителием и заполнен жидкостью. Одна из клеток пузырька впячивается внутрь него в виде вздутой на конце булавы, внутри которой выделяется одна или несколько конкреций углекислой извести. Это статолиты, или слуховые камешки, и они столь же характерны для органов чувства равновесия, как пигмент для органов зрения. Чувствительные клетки пузырька снабжены каждая длинным чувствительным волоском, направленным к находящейся в центре его булаве. Строение волоска сходно со строением книдоциля стрекательных клеток. По функции статоцисты медуз более или менее отвечают функции полукружных каналов уха человека. Новейшие электронно-микроскопические исследования показали, что волоски чувствительных клеток в статоцистах медуз построены по одному типу с чувствительными волосками рецепторных органов более высоко организованных животных вплоть до позвоночных.
В последнее время статоцисты медуз считают не только органами равновесия, но и приспособлениями, стимулирующими сократительные движения краев зонтика: если вырезать у медузы все статоцисты, то она перестанет двигаться.
Медузы раздельнополы. Половые железы (гонады) располагаются у них на нижней стороне зонтика, под радиальными каналами или на ротовом стебельке, и представляют собой кучки половых клеток, лежащих между эктодермой и мезоглеей.
Медузы плавают в толще воды, частью переносимые морскими течениями, частью двигаясь активно при помощи действия мышечных волокон, имеющихся по краю зонтика и в парусе. Одновременным сокращением зонтика и паруса и их последующим расслаблением вода, которая находится в вогнутости зонтика, то выталкивается из него, то пассивно вновь его заполняет. При выталкивании воды животное получает обратный толчок и двигается выпуклой стороной зонтика вперед. Вследствие чередования сокращений и расслаблений зонтика и паруса движение медуз состоит из ряда прерывистых толчков.
Медузы — хищницы. Своими щупальцами они захватывают и убивают различных мелких животных, проглатывают их и переваривают в га-стральной полости.
Половое размножение и развитие.Половые клетки после созревания выходят наружу через небольшие разрывы наружной стенки тела. Опло-

|
|

|

|
Рис. 99. Развитие медузы Aequorea (по Клаусу). Гаструляция происходит посредством иммиграции клеток бластулы (А) на вегетативном полюсе последней (5, В), в результате получается двухслойная личинка планула (Г)
дотворенне и все дальнейшее развитие яиц протекает вне материнского организма. Яйцо испытывает полное равномерное дробление, в результате которого формируется продолговатая бластула (рис. 99). Переход от бластулы к двухслойному состоянию — гаструляция — у громадного
|
большинства кишечнополостных совершается не впячиванием одной половины бластулы в другую, а путем иммиграции. На заднем
полюсе бластулы часть клеток ее стенки начинает поодиночке уходить внутрь личинки, т. е. в блас-тоцель. В конце концов полость заполняется сплошной массой однородных клеток, которые представляют зачаток энтодермы.
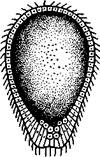
Развившаяся таким образом личинка — паренхимула — напоминает паренхимулу губок (с. ПО). В дальнейшем часть эн-тодермальных клеток личинки дегенерирует и на их месте возникает небольшая полость — зачаток будущей гастральной полости. На этой стадии, характерной для кишечнополостных, личинка называется планулой (рис. 99).
| Рис. 100. Жизненный цикл гидроида Obelia (по Наумову). А — яйцо; 5 — планула; В — колония полипов с бластостилем, на котором видны развивающиеся медузы; Г — отделившаяся медуза |
Планула овальной формы, сплошь покрыта ресничками, при помощи которых она некоторое время свободно плавает, а потом опускается на дно и прикрепляет-
ся к нему слегка расширенным передним концом. В энтодерме осевшей планулы формируется гастральная полость; на конце, противоположном месту прикрепления, прорывается отверстие, сообщающее эту полость с внешней средой, — ротовое отверстие, по краям последнего вырастает венчик щупалец и планула превращается в маленького полипа. Полип растет, почкуется и дает колонию гидроидных полипов.
|
Чередование поколений.Итак, жизнь гидроидных полипов состоит из правильного чередования двух поколений, различных по строению и способу размножения. Одно поколение, полипоидное, ведет сидячий образ жизни и размножается только бесполым способом, производя посредством почкования полипов и медуз. Медузы (второе поколение) отрываются от колоний полипов и переходят к свободному подвижному образу жизни. Они размножаются половым путем и снова дают начало поколению полипов. Такое чередование поколений, размножающихся различным способом, а именно половым и бесполым, называется метагенезом (рис. 100).
| Рис. 101. Tubularia larynx — отдельный гидрант с гонофорами (но Альману): / — ротовой конус. 2 — венчики щупалец, 3 — гонофоры |
Большинство Hydroidea имеет типичное чередование поколений. У некоторых представителей, однако, наблюдается уклонение от намеченной схемы, сказываю-
щесся в частичном подавлении одного из поколений, а именно медузоидного. Образующиеся на колонии медузы перестают отрываться от нее и, оставаясь на месте, т. е. на бласто-стиле, развивают в себе половые клетки. Такие медузы, или медузои-ды, отличаются недоразвитием рта, органов чувств и некоторых других органов. Подавление медузоидного поколения может идти еще дальше, причем медузы постепенно утрачивают характерную форму и превращаются в простые, набитые половыми клетками мешки (гонофоры), сидящие на колонии полипов (рис. 101). Будучи сначала свободнопод-вижным самостоятельным поколением, медузы, таким образом, постепенно становятся как бы половыми органами колонии полипов: интересный пример низведения особи до степени простого органа.
Но иногда изменение в смене поколений идет, по-видимому, в обратном направлении, причем медузоидное поколение получает полное преобладание над полипоидным. По крайней мере, подобно тому как мы встречаем среди гидрозоев формы чисто полипоидные (гидра), не дающие никогда медуз, точно так же есть группа медуз (отр. Trachylida), которые в течение всего жизненного цикла остаются медузами. Они похожи на описанных раньше гидроидных медуз; планула этих медуз не
оседает на дно, но превращается прямо в медузу. Таким образом, здесь чередование поколений исчезает и остается только одно поколение — медузоидное.
Классификация. Отряд 1. Leptolida— формы с различно выраженным чередованием поколений и в полипоидной стадии обычно образующие кустистые или древовидные колонии. В отряде четыре подотряда: Alhecata, Thecaphora, Limnomedusa и Hydrocorallia.
Это самая многочисленная группа гидроидных. Колонии подотрядов Athecata и Thecaphora (см. рис. 95) образуют густые заросли на морском дне, водорослях, ракуш-
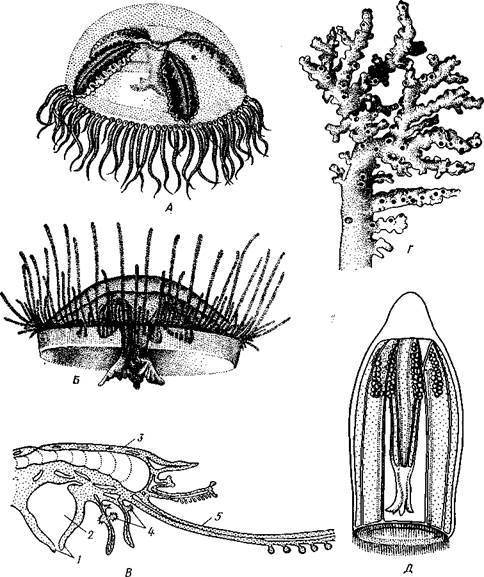
Рис. 102. Различные Hydrozoa. А — ядовитая медуза «Крестовичок» (Gonionemus); £ — пресноводная медуза Craspedacusta; В — схема строения Porpita — представителя отряда Chondrophora; Г—известковая колония Hydrocorallia; Д — трахимедуза Aglantha
(по Наумову, В — из Байера):
/ — ротовое отверстие, 2 — кишечная полость. 3 — хитиноидная скелетная пластинка, 4 — гонофоры,
5 — щупальце
ках, сваях и т. п. Отдельные гидранты очень мелкие, чаще не более 1 мм, только некоторые Athccata, например Tubularia (рис. 101), достигают нескольких сантиметров, а обитающий у берегов Японии одиночный полип Branchiocerianthus—даже до 1 м. Медузы обычно тоже небольшой величины, от 5 мм до 2—3 см в диаметре зонтика, редко бывают крупнее. Большинство представителей отр. Leptolida обитает в море, п пресных водах встречается лишь несколько видов подотряда Limnomedusae, характеризующегося преобладающим развитием медузоидного поколения. Так, в пресноводных водоемах субтропиков и тропиков всех материков, а также в аквариумах с тропическими растениями и рыбками периодически появляются медузы Craspedacusta (рис. 102, Б). К, лимпомеду-зам относится также очень ядовитая морская медуза «крестовичок» (Gonionemus), обитающая у берегов Японского моря и Курильских островов (рис. 102, А). Полипы этих видов едва заметны простым глазом и не образуют колоний.
Представители Hydrocorallia (рис. 102, Г) отличаются массивным известковым скелетом, напоминая этим некоторых коралловых полипов (с. 140). Встречаются в умеренных и тропических морях.
Отряд 2. Chondrophora— крупные одиночные полипоидные формы, обитающие па поверхности воды тропических морей. Тело уплощенное, рот, щупальца и гонофоры обращены вниз (рис. 102, В). На аборалыгой стороне имеется хитиноидная пластинка, соответствующая теке других гидроидных. У Vclella (табл. V, 2) она выступает вверх о виде треугольного паруса.
Отряд 3. Trachylida— чисто мсдузоидные формы, не имеющие смены поколений. Относящаяся сюда Aglantha (рис. 102, Д) встречается по всему Мировому океану, другой представитель — Сипта паразитирует на медузах отряда Leptolida.
Вообще среди кишечнополостных насчитывается очень немного паразитических форм. Особый интерес среди них представляет вид Polypodium hydriforme, систематическое положение которого до сих пор не установлено. Polypodium паразитирует на яйцах осетровых рыб. В икринке хозяина Polypodium имеет вид длинного извитого тяжа— столона, образующего на себе ряд почек со щупальцами. Интересно, что внутри икринки почки Polypodium находятся в вывернутом наизнанку состоянии: эктодерма его обращена внутрь, а энтодерма — наружу к питательному желтку яйца. Позднее почки выворачиваются эктодермой наружу и, когда икра отложена в воду, столон расч издается; образуется 60—90 полипов, которые переходят к свободной жизни, прикрепляясь ко дну. В течение лета полипы размножаются продольным делением. К осени на них образуются половые железы. Дальнейшая судьба полипа и способ заражения хозяина неизвестны.
Отряд 4. Hydrida— одиночные формы, чисто полипоидные, без чередования поколений. Представитель — пресноводная гидра Hydra и немногие другие.
В ископаемом состоянии гидроидные известны, начиная с силура. Главным образом это массивные известковые Hydrocorallia. Имеются также отдельные сведения о нахождении ThecapKiora, уже начиная с кембрия.
ПОДКЛАСС II. СИФОНОФОРЫ (SIPHONOPHORA)
Сифонофоры — свободноплавающие морские животные, встречающиеся преимущественно в теплых морях. Колонии состоят из главного ствола, на котором сидят особи (зооиды) различного строения и физиологического значения. У большей части сифонофор ствол вытянут и особи расположены по всей его длине.
Строение и физиология.Как ствол, так и особи состоят из эктодермы, энтодермы и мезоглеи. Ствол полый, внутренность его занята гастраль-кой полостью, продолжающейся во все особи колонии. Таким образом устанавливается тесная связь между всеми частями колонии. Верхний, слепо замкнутый конец ствола сильно вздут и снабжен глубоким впячи-ванием, наполненным газом. Вздутие называется воздушным пузырем или пневматофором (рис. 103). По краям отверстия, ведущего в полость пузыря, имеется мускул-замыкатель, а эпителий дна пузыря состоит из железистых клеток, способных выделять газ, близкий по составу к воздуху. Воздушный пузырь — сильно видоизмененная медузоидмая особь, есть гидростатический аппарат, позволяющий животному менять удельный вес. Если пузырь наполнен воздухом, то сифонофора плавает у са-
мой поверхности воды. В случае сильного волнения воздух выжимается из пузыря сокращением стенок последнего и животное опускается на
|
глубину. Для поднятия вновь кверху железистые клетки выделяют газ, который благодаря сокращению мускула-замыкателя задерживается в пузыре и раздувает его.
Под пузырем на стволе располагаются плавательные колокола (нектофоры), имеющие форму небольших медуз, но с сильно редуцированным стебельком, щупальцами и органами чувств. Функция плавательных колоколов— передвижение сифонофоры: постоянным сокращением зонтиков они гонят животное воздушным пузырем вперед.
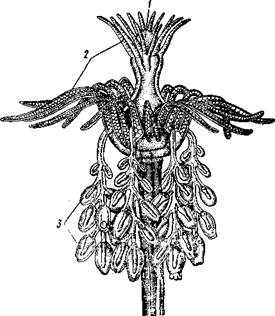
Очень важную часть сифонофоры составляют так называемые кормящие полипы — гастрозоиды. Каждый гастрозоид имеет одно длинное разветвленное щупальце — арканчик. Концы ветвей аркапчика усажены множеством стрекательных клеток, образующих стрекательные батареи. Арканчик захватывает и парализует добычу, которая проглатывается широким ртом гастрозоида и в нем переваривается.

|
Кроме гастрозоидов на стволе сидят еще другие полипообразные особи — пальпопы. Это как бы упрощенные гастрозоиды с неразветвленным простым щупальцем. Функция их неясна, но предполагается, что они имеют выделительное значение. Обязательно имеются и половые медузоиды (гонофоры), построенные по типу медуз (вроде прикреплённых медузоидов на колониях некоторых форм из отр. Leptolida) и содержащие половые железы. На одной колонии бывают два сорта медузоидов — мужские и женские. Расположение особей на теле сифонофор различно, но чаще всего таково: воздушный пузырь один и занимает вершину ствола; под ним располагаются плавательные колокола. Прочие особи сидят на стволе группами, разделенными известными промежутками. Группы называются корми-днями, причем каждая обычно слагается из двух половых медузоидов, гастрозооида и пальпопа. Кроме того, часто кормидий бывает прикрыт особой кроющей пластинкой, представляющей также видоизмененную особь.
Размножение и развитие.При половом раз-
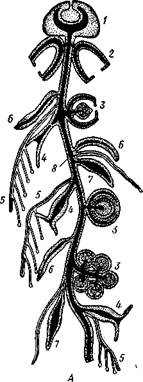
Рис. 103. Схема строения сифонофоры (А) (по Холод-ковскому) и сифонофора Halistemma (Б) (по Деляжу
и Эруару):
Пневматофор, 2 — плавательный колокол (нектофор), 3—половая особь (гонофор), 4—кормящий полип (гастрозоид), 5 — арканчик, 6 — кроющая пластинка, 7 — выделительный полип со щупальцем (пальпон), 8 — ствол колонии
множении из яиц, формирующихся внутри женских мсдузоидов, получаются, как и у других кишечнополостных, личинки планулы; планула вскоре преобразуется в более сложную личинку, на которой отдельные зооиды образуются путем почкования.
|
Экология.Сифонофоры особенно многочисленны в теплых морях. Среди них имеются как мелкие формы, так и виды значительных размеров, у которых длина ствола достигает более 1 м, а длина арканчиков бывает еще больше (у Physalia до Юм). Тело сифонофор прозрачно, но отдельные его части обычно окрашены в яркие цвета. Большинство сифонофор погружено полностью в воду, но у Physalia крупный пневматофор (до 30 см в
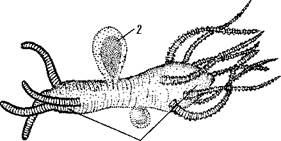
| Рис. 104. Плавающая колония Moerisia с полипо- идными (1) и медузоидными (2) особями (по Наумову) |
длину), находящийся на вершине укороченного ствола, торчит из воды. Эти си-фонофоры уже не могут погружаться в воду, и воздушный пузырь служит им в качестве паруса. Ожоги длинных арканчиков физалии, снабженных мощными стрекательными . батареями, чувствительны даже для человека.
Происхождение сифонофор.При толковании строения сифонофор были высказаны две точки зрения. Одни ученые отстаивали их колониаль-ность, другие считали сифонофору единой особью, а все ее придатки лишь органами сложной особи.
Большинство современных зоологов считают сифонофор происшедшими от колоний полипов, приспособившихся к плавающему образу жизни.
Кроме истории развития, в пользу такого толкования свидетельствует и то, что в настоящее время найдено несколько плавающих колониальных полипов (в том числе один в Каспийском море — Moerisia) (рис. 104). На колониях таких гидроидов развиваются как полипоидные, так и медузоидные особи.
Можно представить себе, что сифонофоры произошли от подобных колониальных Hydroidea в результате возникновения полиморфизма особей и разделения функций между ними. При таком толковании строения сифонофоры воздушный пузырь, плавательные колокола и го-нофоры вполне легко и естественно подводятся под тип строения медуз, а кормящие полипы и пальпоны отвечают видоизмененным полипам.
КЛАСС II. СЦИФОИДНЫЕ МЕДУЗЫ (SCYPHOZOA)
К этому сравнительно небольшому классу (200 видов) относятся медузы, обитающие только в морях. Они значительно крупнее гидромедуз (рис. 105). Кроме того, они легко отличимы от последних по отсутствию паруса. В остальном у сцифомедуз типично медузоидное строение.
Строение и физиология.Тело в виде круглого зонтика или, при вытягивании его по главной оси, высокого колокола (рис. 106). Посреди нижней вогнутой стороны зонтика на конце ротового стебелька помещается четырехугольный рот. Углы рта вытягиваются в 4 желобовидных выро-
| 5—763 |
|
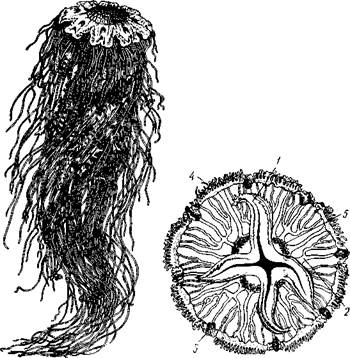
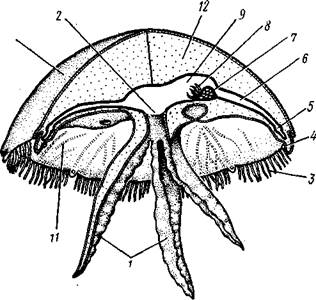
| Рис. 105. Сцифоидные медузы. А — Cyanea capillata (рис. Кондакова); Б —Aurelia aurita (no Кюкенталю) : / — ротовая лопасть, 2 — радиальные каналы гастроваскулярной системы; 3 — ропалии, 4—щупальца по краю зонтика, 5 — гонады |
|
| 10 |
Рис. 106. Схема строения сцифоидной медузы (из Байера)] / — ротовые лопасти, 2—ротовое отверстие, 3 — щупальца, 4 — ро-палий, 5 — кольцевой канал, 6 — радиальный канал, 7 — гонада, 8 — гастральные нити, 9 — желудок, 10 — эксумбрелла, // — субумб-релла, 12 — мезоглея. Эктодерма показана штриховкой, энтодерма — черным
ста — ротовые лопасти, служащие для захвата пищи; у некоторых так называемых корнеротых сцифомедуз (отр. Rhizostomida) ротовые лопас-сти становятся складчатыми и срастаются, так что от ротового отверстия остаются лишь многочисленные мелкие поры, через которые проходит в кишечник пища — мелкие планктонные организмы (см. рис. 109).
|
| J |
Рот ведет в энтодермальный желудок, занимающий центр зонтика и образующий четыре неглубоких карманообразных выпячивания. В желудок вдаются с краев четыре валика с гастральными нитями, которые служат для увеличения всасывающей поверхности энтодермы.
| 1 |
От желудка расходится к краям тела система радиальных каналов. В простейшем случае их всего четыре, у других видов — восемь, у ряда сцифомедуз гастроваскуляр-ная система еще сложнее — состоит из 4 сильно ветвящихся каналов первого порядка, 4 ветвистых каналов второго порядка и 8 неразветвленных каналов третьего порядка. Каналы правильно чередуются в расположении, а своими наружными концами впадают в кольцевой канал, окаймляющий край зонтика.
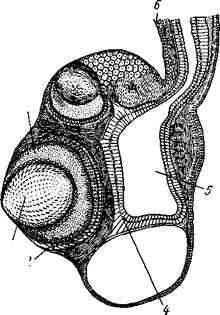
| Рис. 107. Продольный разрез чеоез ропалий медузы Carybdea. Налево разрез прошел через 2 главных глаза ропалия, пустое пространство внизу ропалия — статоцист (статолиты растворились при изготовлении препарата) (по Шевякову): / — хрусталик. 2 — светочувствительный слой ретинальных клеток, 3 — стекловид ное тело. 4 — энтодермальная выстилкапродолжающегося в ропалий радиального канала (5), 6 — эктодерма ропалия |
Край зонтика несет различное число щупалец. Некоторые из щупалец, расположенные у концов главных каналов первого и второго порядков, видоизменяются и превращаются в краевые тельца, или ропа-лии (рис. 107). При этом щупальца укорачиваются и утолщаются, а внутри них развиваются органы зрения « органы равновесия. Каждый ропа-лий чаще содержит один статоцист и несколько глазков разной степени
сложности строения; наряду с глазками, напоминающими таковые гидроидных медуз, здесь имеются и более сложные глаза типа глазного пузыря. Такой глаз получается посредством погружения глазной ямки под эпителий и отшнуровывания ее от поверхности тела, причем ямка замыкается под кожей в глазной пузырь. Кожный эпителий над пузырем остается тонким и прозрачным и называется роговицей. Дно и боковые стенки пузыря состоят, как и у гидромедуз, из пигментных и чувствительных клеток. Часть стенки пузыря, лежащая непосредственно под роговицей, сильно утолщается и образует двояковыпуклый хрусталик. Внутренность пузыря заполнена бесструктурным стекловидным телом, которое выделяется стенками пузыря. Несмотря на сложность строения, глаза, по мнению большинства авторов, служат лишь для различения света и тьмы. В связи с сильным развитием органов чувств центральная нервная система сцифомедуз тоже испытывает усложнение. На протяжении краевого нервного кольца (соответственно 8 ропалиям) возника-
5* 131
ют 8 скоплений нервных клеток, или ганглиев, — первый пример образования значительных нервных узлов.
Размножение и развитие.Медузы раздельнополы. Половые железы образуются из энтодермы нижней поверхности карманов желудка. Созревшие половые клетки выводятся наружу через рот медузы.
После полного и равномерного дробления яйца образуется бластула, а затем типичная мерцательная планула. Она сначала плавает, позднее же прикрепляется передним полюсом к морскому дну. На прежнем заднем, а теперь верхнем полюсе прорывается рот, который ведет внутрь образующейся к этому времени гастральной полости. Вокруг рта развивается венчик щупалец, число которых кратно четырем. Энтодерма гастральной полости дает 4 продольных желудочных валика. В результате этих изменений планула превращается в маленького одиночного полипа— сцифистому (рис. 108), немного похожего на гидру, но устроенного сложнее. Этот полип может путем почкования давать начало другим сцифистомам.
Главный процесс, совершающийся со сцифистомой, — это стробиля-ция: полип делится путем ряда поперечных перетяжек, которые постепенно врезаются с краев в тело полипа, пока из последнего не получится подобие стопки наложенных друг на друга тарелок, соединенных центральным стволом. На этой стадии развития полип называется стробилой.
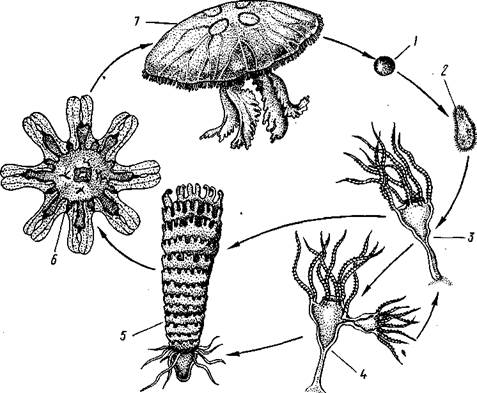
Рис. 108. Развитие сцифомедузы Aurelia:
1 — яйцо, 2— планула, 3—сцифистома, 4 — почкующаяся сцифистома, 5 — стробиляция, 6 — эфира,
7—взрослая медуза (по Ъайеру)
Образовавшиеся в процессе стробиляции диски представляют собой молодых медуз, расположенных вогнутыми сторонами их зонтиков кверху. Медузы постепенно, начиная с верхней, отрываются от сцифистомы, переворачиваются выпуклой стороной кверху и переходят к плавающему образу жизни. Они еще во многом отличаются от взрослых медуз, а потому считаются особой личиночной стадией, или эфирой. Край зонтика эфиры глубоко вырезан в виде 8 лопастей. Превращение во взрослую медузу сопровождается усиленным ростом и состоит главным образом в том, что края зонтика выравниваются, формируется сложная канальная система, появляются краевые щупальца и зачатки гонад.
Таким образом, сцифомедузы обладают ясно выраженным метагенезом, т. е. чередованием полового и бесполого поколений, причем в противоположность Leptolida из гидрозоев у них наиболее развито медузоид-ное поколение.
Экология. Сцифомедузы плавают при помощи сокращений зонтика, число которых может доходить до 100—140 в 1 мин. Некоторые медузы широко распространены; так, Aurelia aurita, обитающая почти во всех умеренных и тропических' морях, заходит и в арктические воды. Другие сцифоидные медузы распространены более ограниченно.
Тело большинства медуз прозрачно, что зависит от большого количества содержащейся в тканях воды (особенно в мезоглее). У многих медуз вода составляет 97,5% общей массы тела.
Размеры сцифомедуз могут быть очень велики: Aurelia aurita достигают обычно 40 см в поперечнике, тогда как Cyanea capillata — иногда почти 2 м в диаметре при длине щупалец 10—15 м. Медузы — хищники. Они питаются различными планктонными беспозвоночными, а иногда и молодью рыб.
Расправленными щупальцами медузы облавливают большую площадь воды. Так, Drymonema, достигая 25 см в поперечнике, облавливает щупальцами круг в 150 м2.
Классификация. Сцифомедузы делятся на 5 отрядов.
Отряд 1. Stauromedusaeсостоит из небольшого числа своеобразных форм, ведущих прикрепленный образ жизни. У них имеется стебелек, которым медуза прикрепляется к субстрату. Край зонтика образует глубокие вырезки, между ними на особых рукооб-разных выростах сидят пучки головчатых щупалец. Жизненный цикл Stauromedusae проходит без чередования поколений. Из планулы непосредственно развивается молодая медузка. Наиболее типичными представителями отряда могут служить Haliclystus (рис. 109, .4) и Lucernaria.
Отряд 2; Cubomedusaeтоже невелик. Его представители имеют обыкновенно четырехгранный высокий зонтик с четырьмя ропалиями (см. рис. 107) и четырьмя простыми (как у Carybdea) либо разветвленными щупальцами. Кубомедузы встречаются в мелководье теплых морей. Некоторые тропические виды, частые у берегов Австралии и Индонезии (например, Chiropsalmus), могут вызывать у человека тяжелые, иногда смертельные «ожоги».
Отряд 3. Coronata.Зонтик делится кольцевой бороздкой на центральный диск и периферическую «корону». Ропалии и щупальца сидят на особых студенистых цоколях. Немногочисленные представители отряда (Atolla, Periphylla) встречаются преимущественно на больших глубинах.
Большинство сцифомедуз относится к двум последним отрядам.
Отряд 4. Semaeostomeae.Характерно наличие дисковидно сплющенного зонтика, несущего по краю многочисленные щупальца. Сюда принадлежат Aurelia aurita (см. рис. 105, Б) и обыкновенная медуза наших северных морей—Cyanea capillata (см. рис. 105, А). Некоторые медузы, особенно Pelagia, способны сильно светиться в темноте.
Отряд 5. Rhizostomida(корнеротые медузы). Обычно это крупные медузы, наибо-ле,е многочисленные в тропических морях. В Черном море встречается представитель этого отряда Rhizostoma pulmo (рис. 109, Б). У корнеротых нет щупалец по краю зонтика. Функция захвата добычи целиком осуществляется ротовыми лопастями. Некоторые корнеротые медузы в странах Азии в соленом виде употребляются в пищу.
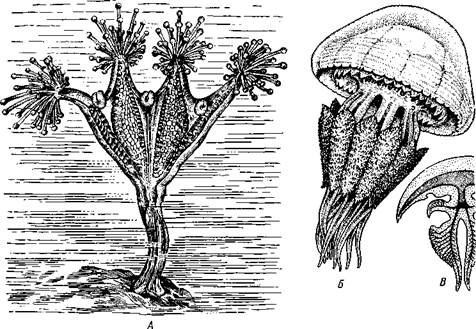
Рис. 109. Представители сцифомедуз. А — сидячая медуза Haliclystus (вид сбоку);
Б — корнеротая медуза Rhizostoma pulmo; В — схема продольного разреза корнеро-
той медузы (по Деляжу и Эреуару)
Мягкость тела и отсутствие скелета у медуз препятствуют хорошему сохранению их в ископаемом состоянии. Однако отдельные отпечатки попадаются, начиная с нижнего Кембрия. Наибольшее количество отпечатков медуз сохранилось в юрском сланце из Золеигофена.
КЛАСС III. КОРАЛЛОВЫЕ ПОЛИПЫ (ANTHOZOA)
Коралловые полипы бывают только полипоидной формы. Они совершенно не обнаруживают чередования поколений. Это морские животные, иногда одиночные, большей частью колониальные, причем колонии могут достигать значительной величины. Это наиболее крупный класс ки-шечнополостных, охватывающий 6000 видов.
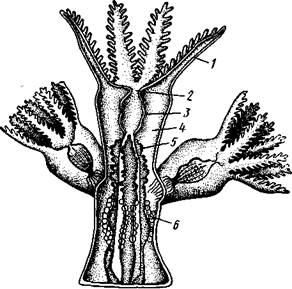
Строение и физиология.Коралловые полипы напоминают гидроидных, но устроены значительно сложнее. Тело отдельной особи имеет форму цилиндра (рис. ПО). Нижний конец одиночных полипов уплощен в подошву, служащую для прикрепления к субстрату, у колониальных форм он соединен со стволом или ветвью колонии. Ротовое отверстие помещается в центре противоположного конца тела. Вокруг рта располагается венчик полых щупалец, число которых у одних полипов (подкл. Octocorallia) равно восьми, в других же (подкл. Hexacorallia) представляет число, кратное шести.
Рот ведет сначала в длинную трубку — глотку, которая свешивается в гастральную полость. Глотка образуется впячиванием стенки ротового диска, вследствие чего выстлана внутри эктодермальным эпителием. Глоточная трубка сплющена в одном направлении, так что просвет глотки получает вид более или менее широкой щели. На одном или обо-
|
| Рис. 110. Схема части колонии восьмилучевого коралла Alcyonium (по Л. и В. Шимкевич): ; — щупальца. 2 — ротовое отверстие. 3 — глотка, 4 — септа, 5 — мезентериальные нити, 6 — яйца |
их концах этой щели расположены ротовые желобки — сифоноглифы (рис. 111), которые несут клетки с очень длинными ресничками. Находясь в непрерывном движении, реснички гонят воду внутрь гастральной полости полипа, тогда как на остальном участке глотки вода выводится из гастральной полости обратно наружу. Таким путем обеспечивается постоянная смена воды, имеющая важное значение для жизни полипа. В кишечную полость поступает свежая, богатая кислородом вода, а у полипов, питающихся мелкими планктонными организмами, и пища. Вода, отдавшая кислород тканям полипа и насыщенная углекислотой, выносится наружу вместе с непереваренными пищевыми остатками.
Собственно гастральная полость выстлана энтодермой и поделена на участки (камеры) особыми радиальными перегородками или септами. Число камер соответствует числу щупалец. Септы состоят из мезоглеи, одетой энтодермой. В верхней части полипа перегородки прирастают
|
| ю |
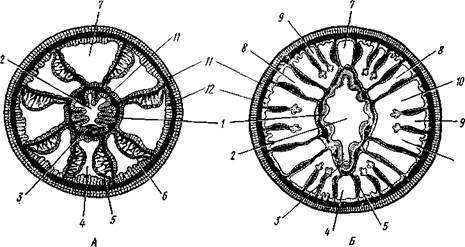
Рис. 111. Поперечные разрезы через восьмилучевой (А) и шестилучевой (Б) коралловые полипы (Л — по Хиксон, Б — по Хайман):
1 — глотка, 2—полость глотки, 3 — сифоноглиф, 4 — вентральная направительная камера, 5 — септа, 6—мускульный валик септы, 7 — дорзальная направительная камера, 8 — внутренние камеры, расположенные между двумя септами первого порядка, 9 ~ внутренние камеры, образующиеся между вторично возникающими септами, 10—промежуточные камеры, 11 — эктодерма,
12 — энтодерма. Мезоглея зачернена

|
одним краем к стенке тела, другим к глотке. В нижней части полипа (ниже глотки) они прикрепляются лишь к стенке тела, вследствие чего центральная часть гастральной полости — желудок — остается неразделенной. Число септ соответствует числу щупалец. Свободные края септ утолщены, они называются мезентериальными нитями (см. рис. ПО). Последние играют важную роль в пищеварении, так как несут железистые энтодермальные клетки, выделяющие пищеварительные ферменты. Гистологическое строение коралловых полипов усложняется по сравнению с гидроидными — вместо тонкой базальной мембраны у них толстый слой мезоглеи, кроме того, вместо эпителиально-мышечных клеток под эктодермой расположен слой обособившихся мускульных клеток (продольных и кольцевых). Произошла дифференциация мускульной ткани. Сильно развитые, энтодермального происхождения мускульные клетки имеются и в септах. Они образуют проходящие вдоль каждой септы (с одной из ее сторон) утолщения — мускульные валики (рис. 111). Подкожное нервное сплетение у коралловых полипов выражено гораздо сильнее, чем у гидроидных.
| Рис. 112. Известковые иглы (спикулы) восьмилучевых кораллов |
|
| Рис. 113. Схема строения шестилуче- вого кораллового полипа: образова ние склеросепт (по Пфуртшеллеру): / — щупальца, 2 — ротовое отверстие. 3 — глотка, 4 — септы, 5—подошвенная пластинка, 6— чашечка, 7 — склеросепты, впячивающие мягкие ткани полипа (8) в гастральную полость |
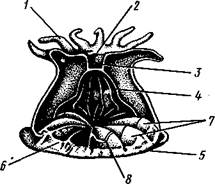
Только одиночные коралловые полипы, да и то не все, лишены скелетных образований. Напротив, у колониальных кораллов есть скелет, чаще всего состоящий из углекислой извести, реже (у части восьми-лучевых кораллов Octocorallia) из рогоподобного вещества. У восьми-лучевых кораллов известковый скелет залегает внутри мезоглеи и в простейшем случае состоит из разбросанных микроскопических известковых игл (рис. 112). Последние, как и у губок, формируются внутри особых клеток — склероблас-тов. У благородного коралла известковых игл (спикул) так много, что большинство их сливается в плотную массу, образуя твердый скелет. У шестилучевых кораллов (рис. 113) известковый скелет устроен иначе. У молодой особи сначала клетками эктодермы выделяется кнаружи подошвенная пластинка, а затем кругом тела полипа скелетная чашечка, или тека. Далее от теки внутрь тела врастают скелетные перегородки (склеросепты), впячивающие перед собой стенку полипа глубоко в его гастральную полость.
У колоний, состоящих из громадного числа особей, чашечки соседних полипов часто сливаются. Итак, у подкл. Octocorallia скелет внутренний (залегает внутри мезоглеи), у подкл. Hexacorallia по происхождению он внешний, так как лежит кнаружи от эктодермы, представляя продукт ее жизнедеятельности.
Склеросепты и настоящие мягкие перегородки имеют вполне определенное число и расположение, служащие важным систематическим признаком.
Восьмилучевые кораллы Octocorallia (см. рис. 111, Л) имеют 8 перегородок, делящих гастральную полость на 8 периферических камер; две из этих камер, приходящиеся против узких краев глоточной трубки, называются направительными. На септах в строго определенном порядке расположены мускульные валики. В результате в полость одной из направительных камер (условно называемую вентральной) обращены два мускульных валика. В другую направительную камеру (дорзальную) мускульные валики не вдаются.
У шестилучевых кораллов расположение септ сложнее (см. рис.'111, Б). Число расположенных попарно перегородок кратно шести, но по меньшей мере их 12. Перегородки возникают не все сразу. Сначала образуются шесть пар перегородок первого порядка, которыми гаст-ральная полость делится на 12 камер. Камеры, лежащие между двумя перегородками одной пары, называются внутренними, расположенные же между перегородками различных пар — промежуточными. Дальнейшие перегородки возникают парами, образуясь в пределах промежуточных камер.
Склеросепты развиваются всегда во внутренних камерах и не препятствуют закладке новых мягких септ, образующихся только в промежуточных камерах.
Физиологические отправления исследованы преимущественно у актиний— одиночных крупных коралловых полипов, лишенных скелета. Актинии в противоположность колониальным формам могут медленно ползать при помощи своей мускулистой подошвы. При раздражении актинии сильно сокращаются, втягивают щупальца и превращаются в небольшой твердый комок. Актинии очень чувствительны, особенно их щупальца.
Крупные актинии — большие хищницы, питаются раками, моллюсками и т. п. Добыча захватывается щупальцами, которые выпускают заряд стрекательных нитей, затем рот раскрывается, часть глотки выворачивается из него в виде широкого мешка и обхватывает пойманное животное.
Пищеварение актиний так же, как у гидр, представляет сочетание внутриклеточного с полостным. •
Размножение и развитие.Коралловые полипы размножаются бесполым и половым путем. Одиночные мягкие актинии иногда размножаются делением, у колониальных форм наблюдается почкование. Коралловые полипы, как правило, раздельнополы. Половые железы формируются в перегородках, между их энтодермой и мезоглеей. При половом размножении сперматозоиды прорывают эпителий септы мужских особей, выходят через рот наружу и через рот же проникают в женские особи, где и происходит оплодотворение яйца. Начальные стадии развития проходят в мезоглее септ. У многих актиний все развитие вплоть до образования полипа происходит в гастральной полости материнского организма. У некоторых Anthozoa оплодотворение наружное.
Развитие коралловых полипов идет сравнительно просто. Яйца испытывают полное равномерное дробление; сначала формируется бластула, потом мерцательная планула, которая плавает, а затем оседает перед-
ним концом на дно и, подобно плануле гидрозоев, превращается в молодого полипа.
Коралловые рифы и роль коралловых полипов в образовании земной коры.В тропических частях Атлантического, Индийского и Тихого океанов распространены рифообразующие (мадрепоровые) кораллы, отличающиеся иногда очень крупными' размерами (более 2 м в диаметре; ветви коралла Асгорога достигают высоты свыше 4 м). У них массивный известковый скелет, они образуют на мелководье густые поселения — коралловые рифы. Наиболее известны рифы побережья Австралии, Индонезии и островов Океании, многие из которых целиком слагаются из кораллового известняка. Дальше всего на север коралловые рифы заходят в Красном море. Самого «цветущего» состояния они достигают там, где зимняя температура воды не опускается ниже 20°С. Вместе с тем эти кораллы имеют ограниченное распределение по вертикали, опускаясь лишь до глубины 50 м. Это связано с тем, что в тканях рифообра-зующих мадрепоровых кораллов живут симбиотические одноклеточные водоросли — зооксантеллы, которые нуждаются в солнечном свете. Роль зооксантелл в жизнедеятельности мадрепоровых кораллов до конца еще не выяснена. Предположение о том, что кораллы способны переваривать эти водоросли, в последнее время не подтвердилось. По-видимому, зооксантеллы необходимы для нормального образования скелета кораллов. Рост скелета резко замедляется в темноте или у кораллов, искусственно лишенных зооксантелл. Все рифообразующие кораллы нуждаются в морской воде с нормальной океанической соленостью, т. е. содержащей около 3,5% солей. В связи с этим они никогда не поселяются в устьях

|
| Рис. 114. Участок кораллового рифа во время отлива (по Гентшелю) |
рек и других опресненных участках моря. Мадрепоровые кораллы нуждаются также в чистой прозрачной воде, достаточно насыщенной кислородом. Коралловый риф служит местом обитания и развития многих морских организмов (pile. 114). Здесь в большом количестве поселяются водоросли, моллюски, черви, ракообразные, иглокожие и представители других групп донных кишечнополостных. В зарослях коралловых рифов обитают многочисленные коралловые рыбки. Все эти животные и растения в совокупности образуют своеобразное сообщество, или биоценоз, кораллового рифа. Часть членов этого сообщества обладает мощным известковым скелетом и наряду с мадрепоровыми кораллами принимают участие в образовании рифа. Другие же находят здесь убежище и питаются за счет рифообразующих организ-мов- Животные кораллового биоценоза имеют причудливую
пеструю расцветку, которая помогает им укрываться на фоне ярко окрашенных колоний мадрепоровых кораллов.
Рифы распадаются на три разновидности: береговые, барьерные и атоллы. Береговые окаймляют берег суши, барьерный риф расположен параллельно берегу, но на некотором от него расстоянии. Особенно знаменит Большой Барьерный риф, тянущийся на протяжении 1400 км вдоль восточного берега Австралии.
Атолл — коралловое кольцо, выдающееся из моря на небольшую высоту, внутри кольца находится озеро морской воды, лагуна. На такой атолл приносятся водой или ветром семена разных растений (в том числе кокосовые орехи) и он превращается в цветущий островок.
Происхождение береговых рифов не требует объяснений. Труднее объяснить возникновение атоллов, так как они со всех сторон окружены большими глубинами, не подходящими для поселения кораллов.
Ч. Дарвин, наблюдавший коралловые рифы и острова во время путешествия на «Бигле», предполагал, что все разновидности рифов возникли из береговых вследствие постепенного опускания дна океана. Если на месте острова, окруженного береговым рифом, морское дно начнет медленно понижаться, то остров станет уходить под уровень воды и уменьшаться в размерах. Кораллы, его окружающие, по мере опускания рифа будут надстраивать его кверху, так как живут лишь на небольших глубинах, более же глубокие части рифа начнут отмирать. Таким образом, в то время как сам остров будет опускаться, риф, его окаймляющий, останется у поверхности воды, постепенно отделится от сократившегося в размерах острова и превратится в барьерный. При полном опускании острова на его месте остается мелководная лагуна, а риф превращается в атолл.
Другими учеными (Агассиц и др.) возникновение барьерных и атолловых рифов объясняется иначе. Они считают, что кораллы покрывают самые вершины находящихся под водой горных кряжей и пиков. Эти горы сначала выдавались над водой, но потом были настолько разрушены атмосферными влияниями, что ушли под уровень моря, а на месте самых высоких точек их образовались мелководные участки моря, очень удобные для поселения кораллов. Бурение атоллов показало, однако, что на несколько сотен метров в глубину их основание состоит из отмершего кораллового полипняка. Это подкрепляет не взгляды Агассица, а теорию, высказанную Дарвином.
Некоторые авторы (Меррей) принимают, что атоллы и барьерные рифы возникают вследствие растворяющего известь действия морской воды. Наружные, обращенные к открытому океану части рифа омываются водой, богатой солями извести, что способствует росту полипняков, внутренние же части рифа пользуются водой, которая обеднена известью, но зато обогащена углекислотой — продуктом дыхания коралловых полипов, способствующей растворению извести. Вследствие этого отмирающие внутренние части рифа постепенно растворяются морской водой, тогда как периферические продолжают надстраиваться. Однако в результате последующих гидрохимических исследований моря теория Меррея не получила подтверждения. Оказалось, что углекислота, содержащаяся в воде лагуны, так же, как и по внешнему краю рифа, в условиях высокой температуры тропиков известь не растворяет. Таким образом, ни одна из более новых теорий происхождения коралловых рифов и островов не опровергла основных положений Дарвина.
Что касается способа происхождения надводной части рифа, то все рифы сначала подводные, затем постепенно море обламывает краевые части рифа и обломки выкидывает на поверхность. Слой обломков рас-
тет, промежутки между крупными полипняками заполняются мелким обломочным материалом и, наконец, на месте бывшего подводного рифа получается слой слежавшейся известковой массы до 4 м и более высоты.
Мощная созидательная деятельность полипов объясняется отчасти их огромным количеством, отчасти легкостью процесса усвоения извести из морской воды в тропических водах.
Коралловые рифы существовали, начиная с древнейших геологических эпох, но состав животных-рифообразователей менялся. Рифы кайнозоя и мезозоя были построены из кораллов, сходных с современными. В палеозое строителями рифов были вымершие коралловые полипы из подклассов Rugosa и Tabulata. Кроме того, большое участие в создании рифов принимали Strornatoporoidea — вымершие представители Hydro-zoa, близкие к современному подотр. Hydrocorallia.
Классификация.К классу Anthozoa относятся два современных и три целиком вымерших подкласса.
ПОДКЛАСС I. ВОСЬМИЛУЧЕВЫЕ КОРАЛЛЫ (OCTOCORALLIA)
Представители подкласса имеют 8 щупалец и 8 септ, скелет Octoco-rallia развивается в мезоглее (рис. 115). Объединяет отряды: альционарии (Alcyonaria), горгонарии, или роговые кораллы (Gorgonaria), и морские перья (Pennatularia). К альционариям принадлежат наиболее просто устроенные коралловые полипы. Скелет их состоит из разбросанных в мезоглее спикул, никогда не сливающихся в осевой стержень. Колонии альционарии образуют густые заросли на небольших глубинах тропических морей, но встречаются также в умеренных и полярных водах. В северных и дальневосточных морях СССР наиболее обычна разветвленная
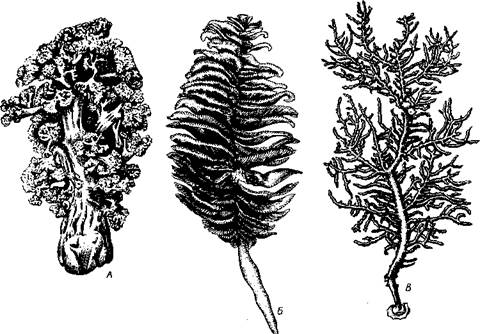
Рис. 115. Различные восьмилучевые кораллы. А — альционария Gersemia; Б — морское перо Pennatula; В — колония рогового коралла Leptogorgia
Gersemia. У Gorgon aria кроме спикул по оси древовидной колонии залегает роговой стержень, иногда частично или полностью обызвествлен-ный. Некоторые роговые кораллы замечательны тем, что их скелет содержит значительное количество иода. Этим, вероятно, объясняется применение их в средние века в качестве лекарственного средства. Особое значение имеет благородный коралл (Corallium rubrum); встречается на глубине свыше 50 м в Средиземном море. Известковый скелет благородного коралла, окрашенный в розовый или кир.пично-красный цвет, служит для изготовления украшений и мелких поделок. Изящные морские перья с правильным двухрядным расположением полипов на прямом стволе оранжевой, розовой или фиолетовой колонии относятся к отряду Pennatularia. Главный ствол такой колонии образован первичным полипом; в толще ствола нередко залегает роговой опорный стержень. Колонии морских перьев могут внедряться своим основанием в песчаный или илистый грунт. Некоторые из них обладают способностью к свечению.
ПОДКЛАСС II. ШЕСТИЛУЧЕВЫЕ КОРАЛЛЫ (HEXACORALLIA)

|
Шестилучевые кораллы делятся на пять отрядов, из которых наиболее обширны отряды актиний (Actiniaria) и мадрепоровых (Madrepora-ria). Актинии — одиночные полипы, способные медленно ползать при помощи подошвы. Это лишенные скелета крупные полипы (иногда свыше 60 см в диаметре), отличающиеся правильностью формы и красотой расцветки. Некоторые актинии (Sagarlia и др.) живут в симбиозе с раками-отшельниками, на раковинах которых они поселяются (рис. 116). При этом рак служит для актиний средством передвижения, тогда как близкое соседство актиний, вооруженных стрекательными капсулами, защищает рака от нападения хищников. Вырастая и переселяясь в более крупную раковину, рак пересаживает актиний клешнями с прежней раковины на новую. Мадрепоровые кораллы (Mad-reporaria) образуют колонии с массивным наружным извест- Рис' 116' Актиния на Раковине, занятой раком-
FJ .. отшельником (по Макарову)
ковым скелетом. Мадрепоро
вые кораллы — основные обра-
зователи коралловых рифов; известны, начиная с триаса. Кроме того, имеются отряды одиночных (Ceriantharia) и колониальных (Zoantharia и Antipatharia) полипов.
ПОДКЛАСС III. ЧЕТЫРЕХЛУЧЕВЫЕ КОРАЛЛЫ (TETRACORALLIA, ИЛИ RUGOSA)
Это вымершие палеозойские, большей частью одиночные (рис. 117, А), реже колониальные кораллы; чаще имеют вид расширенного к верхнему концу бокала или рога. В молодом возрасте Tetracorallia обнаруживают, судя по их скелетам, закладку
|

|

|
Рис. 117. Ископаемые кораллы. А—одиночный четырехлучевой коралл Omphiutna (из Деляжа); Б — колония кораллов Tabulata (из Друскина и Обручевой); В — колония
Heliolitidae (из Байера): / — общий вид, 2 — участок колонии
6 первичных перегородок, что говорит в пользу их родства с подкл. Hexacorallia. Следующие склеросепты закладываются не во всех шести, а только в четырех секторах. Отсюда и название этих кораллов — четырехлучевые. Второе наименование — Rugosa — связано с характерными кольцевыми морщинами на наружной поверхности скелета. Четырехлучевые кораллы распространены с ордовика до пермской эпохи, но главного развития достигают в каменноугольную эпоху, когда за их счет образовывались значительные коралловые рифы,
ПОДКЛАСС IV. ТАБУЛЯТЫ (TABULATA)
Вымершие колониальные (рис. 117, Б), реже одиночные кораллы с полипами незначительных размеров. Число склеросепт обычно кратно 6, но, как правило, они недоразвиты или имеют вид расположенных рядами шипиков. Древнейшие табуляты известны с кембрия, но наиболее широкое распространение получили в ордовике, силуре и девоне, участвуя в образовании коралловых рифов. К началу мезозоя табуляты полностью вымерли.
ПОДКЛАСС V. ГЕЛИОЛИТИДЫ (HELIOLITIDAE)
Большая группа вымерших палеозойских кораллов, исключительно колониальных. Они характеризуются наличием 12 склеросепт. Гелиолитиды были широко распространены и поэтому имеют большое значение как руководящие формы при определении возраста палеозойских отложений.
Филогения типа Coelenterata
Низшими из кишечнополостных являются, без сомнения, гидрозои (Hydrozoa). Это доказывается тем, что у них тело представляет собой двухслойный мешок, оба слоя которого соприкасаются по краям рта. Исходной формой гидрозоев следует, по-видимому, считать морских гидроидных полипов, которые уже вторично дали начало медузам, отличающимся гораздо более сложной организацией. Появление медуз имело прогрессивное значение, так как они играли большую роль в расселении вида. Однако у современных Hydrozoa в ряде случаев произошла вторичная утрата медузоидного поколения.
Пресноводные Hydrida не могут рассматриваться как примитивные в филогенетическом смысле формы, так как особенности их строения и
жизненного цикла, по-видимому, вторично изменены в связи с переходом к пресноводному образу жизни.
Среди гидрозоев сифонофоры представляют, вероятно, группу более позднего происхождения, у -которой произошла значительная и разно-направленная специализация отдельных особей колоний, в результате чего получились полиморфные колонии.
В процессе эволюции Scyphozoa полипоидная стадия (сцифистома) усложнилась незначительно, тогда как сцифомедузы достигли гораздо более высокой степени сложности, что связано с их свободноплаваюшим образом жизни. Эволюция Anthozoa, жизненный цикл которых проходил без метагенеза, шла в направлении приспособления к сидячему образу жизни, для которого очень характерно размножение почкованием и образование колоний. Этот класс кишечнополостных проделал значительную прогрессивную
Дата добавления: 2015-01-24; просмотров: 2207;