Половые клетки, лежащие в паренхиме
5 мм) животные (см. рис. 134, В) с телом большею частью лишь слабо приплюснутым, почти цилиндрическим или веретеновидным. Многие из них хорошо плавают в воде при помощи биения покрывающих тело ресниц. Характерную черту организации образует прямой, слепо замкнутый сзади кишечник.
Ротовое отверстие обычно расположено у переднего конца тела. Над глоткой помещается небольшой мозговой ганглий, от которого идут назад 3, 2 или даже 1 пара
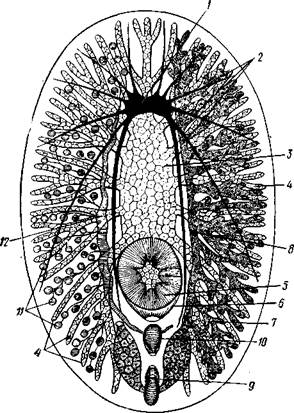
Рис. 135. Организация Polycladida (по Граффу):
/ — мозговой ганглий, 2 — продольные нервные стволы, 3 — средняя часть кишечника, 4 — боковые ветви кишечника, 5 — глотка. 6 — глоточный карман, 7 — яичники, 8 — яйцевод, 9 — женское половое отверстие, 10 — совокупительный орган. 11 — семенники, 12 — семяпровод. На рисунке слева удалены яичники, справа — семенники
нервных стволов. Из органов чувств большей частью имеется пара очень просто устроенных глазков в переднем конце тела. Выделительную систему образует 1 пара каналов с двумя отверстиями. В половой системе характерно массивное строение гонад, которые чаще всего состоят из одной пары семенников, яичников и желточников. Расположение мужского и женского половых отверстий весьма варьирует. При этом в ряде случаев женская половая система помимо протока, служащего для выделения яиц, имеет другой проток специально для совокупления — влагалище. Мы даем столь подробное описание этого отряда ввиду того, что этой группе ресничных червей придается в настоящее время большое филогенетическое значение как исходной группы для других классов червей, с чем мы ознакомимся позже.
Прямокишечные встречаются в пресных водах, морях и на суше (во мху). Имеются и паразитические формы, поселяющиеся на моллюсках и других беспозвоночных.
Отряд7. Темноцефалы (Themnocephalida). Эктопаразиты тропических пресноводных ракообразных, моллюсков, водных черепах. Мелкие, несколько миллиметров длиной, уплощенные с вытянутым телом черви, обладающие спереди пучком щупалец (рис. 136, А), а сзади прикрепительной присоской с клейкими железами. Покровы лишены
ресничек. В организации темноцефал есть много общего с прямокишечными турбелляриями, тем не менее их систематическое положение спорно. Ранее их помещали среди сосальщиков, позднее выделяли в самостоятельный класс. Сейчас большинство исследователей считают темноцефал турбелляриями, сильно измененными паразитическим образом жизни.
Отряд 8. Удонеллиды (Udonellida).Мелкие морские турбеллярии, тело которых имеет лишь несколько миллиметров в длину и несет на заднем конце большую присоску (рис. 136, 5), снабженную клейкими железами (приспособление к паразитическому образу жизни). Удонеллиды поселяются на теле рачков, которые, в свою очередь, являются паразитами рыб. Ротовое отверстие ведет в глотку и кольцевидный кишечник, огибающий комплекс половых желез. Развитие прямое. Систематическое положение удонеллид спорно.
|
| 10 |
| /4 |
| яичн мозговой ганг зисты |
| , — 11 ~ глотка, 12 |
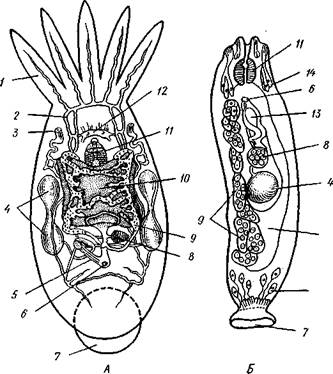
Рис. 136. Паразитические турбеллярии. А — организация
Themnocephala (по Бреслау); Б — удонеллиды (Udonellida):
1 — пучок щупалец. 2 — выделительный канал, 3 ~ экскреторное от
верстие, 4 — семенники. 5 — копулятивный орган, 6 — половое от
верстие, 7 — присоска, 8 — яичник, 9 — желточник, 10 — кишечник,
~ — глий, 13 — зрелое яйцо, 14 — желе
ые клетки
Происхождение турбеллярии.При рассмотрении турбеллярии мы сталкиваемся с важными вопросами возникновения билатеральной симметрии и происхождения всего типа плоских червей. По вопросу о происхождении турбеллярии наибольшее распространение получили две теории. Долгое время популярностью пользовалась теория Ланга, которая основывается на сходстве организации многоветвистых турбеллярии и гребневиков. И те и другие движутся при помощи ресничек, у обеих групп пищеварительная система устроена по общему типу, состоит из глотки, желудка и отходящих от него слепых ветвей. Подобно гребневикам, некоторые турбеллярии имеют расположенный над мозгом орган
| 6—763 |
чувства равновесия (статоцист). Наконец, в развитии гребневиков имеются наметки на закладку третьего зародышевого листка — мезодермы, который уже отчетливо выражен у турбеллярий. Сходство турбеллярий и гребневиков казалось очень убедительным при сравнении поликладид с видом Coeloplana metschnikovi, который был описан русским зоологом А. О. Ковалевским. Coeloplana, как и турбеллярий, сплошь покрыта мелкими ресничками, лишена гребных пластинок, листовидно сплющена и может не только плавать, но и ползать по субстрату; кишечник ее сильно разветвлен. По мнению Ланга, эти гребневики обнаруживают известный переход к организации, характерной для отр. Polycladida, и дают ключ к пониманию происхождения плоских червей. Однако теория Ланга во многих отношениях уязвима. В основу филогенетического древа турбеллярий она ставит сложно устроенных поликладид, в то же время примитивных Acoela рассматривает как вторично упрощенных животных. Основываясь на внешнем сходстве и на строении кишечника обеих групп, теория оставляет без внимания особенности строения половой системы и других органов турбеллярий. Кроме того, данные по эмбриональному развитию поликладид и гребневиков, сравнительный анализ соответствия осей тела этих животных показали несостоятельность теории Ланга.
Вторая теория происхождения турбеллярий, выдвинутая Граффом и впоследствии модернизированная и дополненная известным советским зоологом В. Н. Беклемишевым, получила более широкое распространение. В этом случае исходным считается отряд Acoela, характеризующийся множеством примитивных признаков организации. Отсутствие кишечника позволяет сравнивать Acoela с планулой (или точнее с паренхимулой) кишечнополостных и высказать предположение о происхождении турбеллярий от каких-то сходных с ней предков. Это могли быть личинки древних кишечнополостных, начавшие размножаться еще до превращения их во взрослую форму (способность к размножению на стадии личинки — явление распространенное и называется неотенией). Дальнейшее усложнение таких неотенических личинок постепенно привело к появлению черт строения, характерных для низших турбеллярий. У паренхимул, как известно, нет кишечной полости, и легко представить себе возможность превращения их энтодермы в пищеварительную паренхиму Acoela.
Однако и эта теория, на наш взгляд, не лишена недостатков. Дело в том, что личинка кишечнополостных представляет собой специализированную расселительную личинку, вторично утратившую способность к питанию. Развивающийся из нее полип начинает заглатывать пищу лишь после прикрепления личинки к субстрату и окончательного формирования гастральной полости и ротового отверстия. Приняв теорию Граффа, приходится допустить, что паренхимула не только начала размножаться неотенически, но и вернулась к более примитивному типу питания, который, по-видимому, был свойствен фагоцителлообразным предкам многоклеточных. Эти натяжки отпадают сами собой, если мы примем, что бескишечные турбеллярий ведут свое начало не от неотенических личинок кишечнополостных, а непосредственно от фагоцителлы (с. 93) или подобных ей примитивных многоклеточных организмов.
По-видимому, филогенетические взаимоотношения между низшими группами Metazoa (кишечнополостными и бескишечными ресничными червями) можно представить следующим образом. И тип Coelenterata, и примитивные ресничные черви произошли от фагоцителлообразных предков. В первом случае предковые формы прикреплялись к субстрату, теряя при этом способность активно передвигаться. За счет расхожде-
ния клеток фагоцитобласта (с. 93) у них формируется гастральная полость, выстланная энтодермой, и прорывается ротовое отверстие. Таким образом, мы приходим к исходному полипоидному организму, характерному для кишечнополостных (с. 116), и только у их расселительной личинки сохраняются отчетливо выраженные признаки предковых форм (с. 577).
Во втором случае первичные многоклеточные организмы не прикреплялись к субстрату, а ложились на него боковой поверхностью тела, сохраняя при этом способность к передвижению. Появление постоянного ротового отверстия, превращение фагоцитобласта в пищеварительную паренхиму и развитие примитивных форм полового размножения приводят нас к ацелообразным предкам двусторонне-симметричных животных, которые, по-видимому, и дали начало настоящим бескишечным тур-белляриям. Отсутствие у последних специальных органов выделения, оформленных (одетых оболочкой) гонад и половых протоков, выведение половых продуктов путем разрыва стенки тела или через рот — все эти и другие примитивные особенности Acoela могут найти свое объяснение в исходной простоте организации их фагоцителлообразного предка.
Переход первичных форм к придонному ползающему образу жизни привел к изменению лучистой симметрии тела на билатеральную. У прикрепленных и пассивно плавающих планктонных животных все стороны тела вокруг главной оси находятся в одинаковом отношении к захвату пищи, раздражению внешними стимулами и т. д. Когда же подвижные фагоцителлообразные организмы оказались на дне, эти отношения изменились. Сторона тела, соприкасающаяся с субстратом, стала брюшной, а конец, направленный при ползании вперед, оказался в особых условиях в смысле получения раздражений извне, что послужило причиной развития морфологических отличий спинной и брюшной сторон тела, а на переднем конце постепенно сконцентрировались нервные элементы и образовался мозг. Рот, который у исходных форм, так же как и у планул кишечнополостных, прорывался на заднем конце тела, с переходом предков турбеллярий к ползанию, вероятно, сместился на брюшную поверхность. Это должно было облегчить подбирание пищевых частиц с субстрата. Однако дифференцировка переднего конца тела, на котором оказались сосредоточены органы чувств, сделала более выгодным (в смысле поисков пищи, а с переходом к хищничеству — и захвата добычи) переднее положение ротового отверстия. Очевидно, в ходе эволюции турбеллярий происходил процесс постепенного смещения рта, сохранившего свое положение на брюшной стороне, к переднему концу тела. Отражение этого процесса можно видеть в разнообразии положения ротового отверстия у разных групп современных турбеллярий. Совокупностью всех перечисленных изменений в конце концов определилось возникновение у предков ресничных червей билатеральной симметрии.
В пределах самого класса турбеллярий эволюция пошла по пути развития нервной системы и органов выделения, совершенствования половой системы и других систем органов. Большую роль в этих процессах играло явление олигомеризации (уменьшалось исходно большое число нервных стволов, выделительных каналов, глаз, гонад и т. д.).
КЛАСС II. СОСАЛЬЩИКИ (TREMATODA)
Класс сосальщиков состоит целиком из паразитов, поселяющихся во внутренних органах беспозвоночных и позвоночных животных. К сосальщикам относится около 4000 видов.
6* 163
Организация сосальщиков крайне напоминает таковую турбеллярий, так что при описании некоторых систем органов мы ограничимся лишь краткими дополнениями (рис. 137).
Размеры сосальщиков большей частью измеряются миллиметрами, но иногда черви бывают крупнее. Так, печеночная двуустка достигает 5 см. Наиболее велики некоторые сосальщики из рыб, например представители сем. Didymozoidae, длина тела которых достигает 1,5 м.
Строение.Форма тела чаще всего листовидная. Характерно наличие присосок, от которых происходит и название самого класса. Это блюд-цеобразные ямки, обведенные мускульным валиком, содержащим сложную систему мышечных волокон. Действием этих мышц полость присоски может уменьшаться и увеличиваться.
Обычно имеется одна присоска на переднем конце тела (в глубине ее помещается рот) и одна присоска на брюшной стороне — соответственно ротовая и брюшная присоски. Это органы прикрепления,
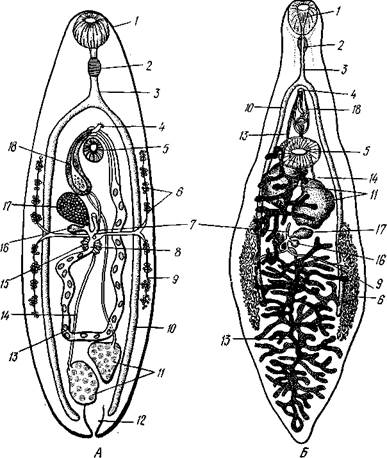
Рис. 137. Организация'дигенетических сосальщиков. А — схема строения сосальщика (по Смиту); Б — организация ланцетовидной двуустки (Dic-
rocoelium dendriticum) (по Кестнеру):
Ротовая присоска, 2 — глотка, 3 — пищевод, 4 — половое отверстие, 5 — брюшная присоска, 6 — желточники, 7 — лауреров канал, 8 — оотип, 9 — желточный проток, 10 ~ ветви кишечника, // — семенники, 12 — экскреторный пузырь, 13 — матка, 14 — семяпроводы, 15 — скорлуповые железы, 16 — семяприемник, 17 — яичник, 18 — совокупительный орган
с помощью которых сосальщики удерживаются в теле животного хозяина.
|
Покровы трематод — тегумент — представлены погруженным эпите
лием (с. 150), но в отличие от такового турбеллярий лишены ресничек.
Эта особенность, по-видимому, связана с паразитическим образом жиз
ни трематод. Наружная часть покро
вов представляет безъядерную ци- . .д
топлазматическую пластинку, содер-
жащую многочисленные митохонд
рии и вакуоли. При помощи цито-
плазматических тяжей этот слой со-
единяется с погруженными в парен
химу участками цитоплазмы, в ко
торых помещаются ядра (рис. 138).
В цитоплазматической пластинке
нередко имеются кутикулярные ши-
пики — дополнительные органы при
крепления паразитов. Этот слой
подостлан базальной мембраной, за
которой следуют кольцевые и про
дольные мышцы. Мускулатура и
паренхима устроены так же, как у
ресничных червей. Сосальщики
сравнительно мало подвижны.
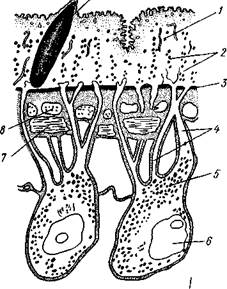
| Рис.138. Схема строения покровов трематод по данным электронной микроскопии (по Тредгольду): 1 — наружная безъядерная цитоплазматиче-ская пластинка, 2 — митохондрии, 3 — базаль-ная мембрана, 4 — цитоплазматические тяжи, соединяющие наружную и погруженную части эпителия, 5 — погруженные участки цитоплазмы с ядрами (6), 7 — продольные мышцы, S — кольцевые мышцы, 9~ кутикулярный ши-пик (продольный срез) |
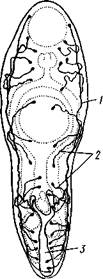
Пищеварительная система. Рот, находящийся на переднем конце тела, ведет в мускулистую эктодср-мальную глотку, а та продолжается в узкий пищевод. Энтодермальная средняя кишка чаще всего слагается из двух ветвей, отходящих от пищевода и тянущихся по бокам тела кзади, где обе ветви заканчиваются слепо. У трематод, отличающихся крупными размерами, например у печеночной двуустки (Fasciola hepa-tica), кишечник многократно ветвится (рис. 139, Б). Это облегчает процесс распределения продуктов пищеварения в паренхиматозном теле животного (с. 152).
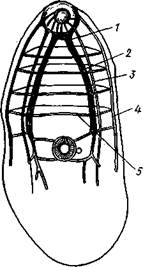
Нервная система (рис. 140) состоит из парного мозгового ганглия, от которого вперед отходят нервы к переднему концу тела и ротовой присоске, а назад — три пары продольных нервных стволов. Сильнее всего развиты брюшные стволы, достигающие значительной толщины. Все продольные стволы соединены кольцевыми перемычками, образуя нервную систему типа ортогона.
Органы чувств развиты крайне слабо, что определяется паразитизмом этих червей. У личинок сосальщиков, некоторое время свободно плавающих в воде, нередко имеются небольшие глазки (одна или две пары), устроенные по типу таковых турбеллярий. Кожные рецепторы (сенсиллы), построенные так же, как у турбеллярий, развиты преимущественно у свободных личинок.
Выделительная система (рис. 141) протонефридиального типа и состоит обычно из пары главных собирательных каналов, от которых расходятся в разные стороны многочисленные веточки, заканчивающиеся
звездчатыми клетками с мерцательным пламенем. Главные каналы открываются на заднем конце тела в общий резервуар — мочевой пузырь, а последний — выделительным отверстием наружу.
Половая система в деталях варьирует, так что удобнее подробно разобрать один частный случай, например половой аппарат ланцетовидной двуустки (Dicrocoelium dendriticum; см. рис. 137, Б). Мужской отдел образован двумя (у громадного большинства сосальщиков) округлыми семенниками, лежащими позади брюшной присоски. От них идут вперед два семяпровода, сливающихся впереди брюшной присоски и образующих извитой семяизвергательный канал; последний пронизывает мускулистый совокупительный орган, способный выпячиваться из тела наружу. Он направлен своим концом в особое мешковидное впя-чивание стенки тела — половую клоаку.
Яичник всегда один и в данном случае лежит несколько позади семенников. Короткий яйцевод ведет от него к небольшому мешочку — оотипу,
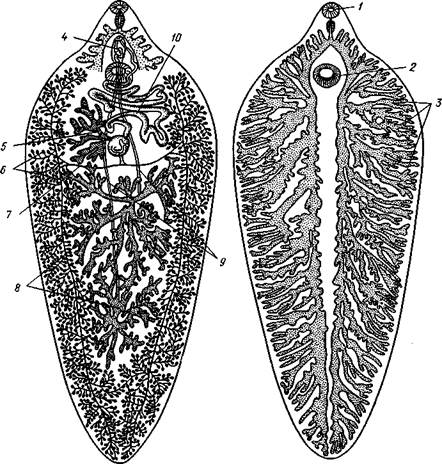
Рис. 139. Печеночная двуустка Fasciola hepatica (из Чендлера). А — строение
половой системы; Б — пищеварительная система:
1 — ротовая присоска, 2 — брюшная присоска, 3 — разветвленный кишечник, 4 — совокупительный орган, 5 — яичник, 6 — желточники, 7 — желточные протоки, 8 — семенники,
9 — семяпровод, 10 — матка
в который впадает большинство протоков женской половой системы. По бокам тела лежат гроздевидные желточники с заключенным в них питательным материалом; два протока желточников сходятся поперек тела к оотипу и впадают в него; от оотипа же отходит длинный извитой канал — матка, идущая сначала назад, потом поворачивающаяся обрат-
|
|
Рис. 140. Нервная система трематоды Microphal-lus (из Белопольской, изменено) :
Дата добавления: 2015-01-24; просмотров: 2325;