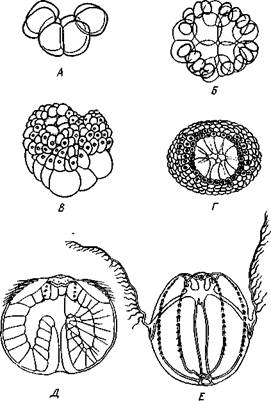
Колпачок (колокол) из слившихся ресничек, 2 — дужки, 3—статолит, 4 — мерцательные бороздки
от поверхности щупальца, но остаются соединенными с ним посредством эластичной спиральной нити, которая, как пружина, амортизирует рывки захваченного животного. Пойманная щупальцами добыча захватывается подвижными краями широкого рта.
Нервная система гребневиков состоит из поверхностного сплетения нервных клеток, которые под рядами гребных пластинок и мерцательными шнурами образуют более плотные тяжи, идущие к аборальному полюсу. Здесь расположен своеобразный орган чувств, получивший название аборального органа (рис. 120). Основу его составляет утолщение эктодермы в виде подушечки, прикрытой прозрачным колпаком, образующимся из венчика слипшихся между собой длинных ресничек. Внутри колпака над поверхностью подушечки расположены 4 крючковидные дужки, образованные длинными слившимися жгутиками, отходящими от рецепторных клеток, связанных с аборальным органом. На дужках подвешен статолит, состоящий из склеенных между собой зерен фосфорнокислого кальция. Основания дужек сообщаются при помощи мерцательных жгутов с рядами гребных пластинок. Хотя весь этот аппарат трактуется как орган равновесия, он служит также и для регуляции движения. У животного с удаленным аборальным органом движение гребных пластинок нарушается.
Мезоглея гребневиков хорошо развита, прозрачна и водяниста, как у медуз. Гребневики не отличаются яркостью окраски, иногда имеют слегка розовый оттенок. Бьющие, прозрачные гребные пластинки переливаются на общем фоне тела всеми цветами радуги.
Размножение и развитие.Гребневики гермафродиты, причем половые клетки их дифференцируются в энтодерме. По бокам каждого меридионального канала пищеварительной системы залегает с одной стороны колбасовидный яичник, с другой — такой же семенник.
Зрелые половые клетки выходят посредством прорыва разделяющей их стенки в просвет меридиональных каналов, а оттуда через рот наружу, где и происходит оплодотворение.
Жизненный цикл гребневиков простой, без метаморфоза. Дробление
полное, но не равномерное, один полюс зародыша состоит из быстро
|
| Рис. 121. Эмбриональное развитие гребневика Во-linopsis (из Байера). А — ранняя стадия дробле-вия; Б — образование микромеров; В, Г — стадии гасгруляции; Д — продольный разрез молодой личинки; Е — сформированная личинка |
дробящихся мелких микромеров, другой — из медленно делящихся макромеров (рис. 121). Микромеры дают эктодерму, макромеры — энтодерму. Микромеры обрастают книзу массу макромеров, которая к тому времени обнаруживает на нижнем полюсе впячивание — первичный рот и зачаток гаст-ральной полости. Получается гаструла. По краям бла-стопора эктодерма впячивается и образует глотку. В глубине гастральной полости часть клеток энтодермы уходит в промежуток между эктодермой и энтодермой (т. е. в остаток полости бластулы) и дает там крестообразную группу клеток. Из этих клеток впоследствии формируются клеточные элементы мезоглеи и мускульные оси щупалец. Интересно, что здесь мы впервые видим во время развития обособленные зачатки специального третьего клеточного зародышевого пласта — мезодермы. Этим слабым зачатком является вышеупомянутая крестообразная клеточная пластинка.
Классификация.Ctenophora делятся на два подкласса: Tentaculata и Atentaculata. Первые в течение всей жизни или только на ранних стадиях развития имеют щупальца, вторые лишены щупалец на всех стадиях. Большинство видов относится к щупальцевым (Tentaculata). Некоторые из них имеют сильно измененную форму тела, которое вытягивается в поперечном направлении (в глоточной плоскости) в ленту до 1,5 м длины; таков «Венерин пояс» — Cestus veneris (рис. 122,5). Большой интерес представляют Platyctenidea. Эти гребневики (рис. 122, В) сплющены по направлению главной оси и не только плавают, но могут и ползать по субстрату на уплощенной ротовой стороне. При переходе от плавающего к ползающему образу жизни организация гребневиков претерпевает глубокие изменения. Исключение среди них представляет арктическая форма Tjalfielta (рис. 122,Г), которая ведет сидячий образ жизни. Молодь ее имеет вид типичных гребневиков. Молодое животное садится ртом на субстрат, средняя часть рта зарастает, а его концы вытягиваются кверху в трубки, служащие для захвата пищи. К бесщупальцевым гребневикам относится обычный в северных и дальневосточных морях Вегое сисщтз (рис. 122, Д). Питается он другими гребневиками, заглатывая их широко раскрывающимся ртом.
Филогения типа Ctenophora.Гребневики обладают некоторыми при-митивными чертами, например движением при помощи измененных
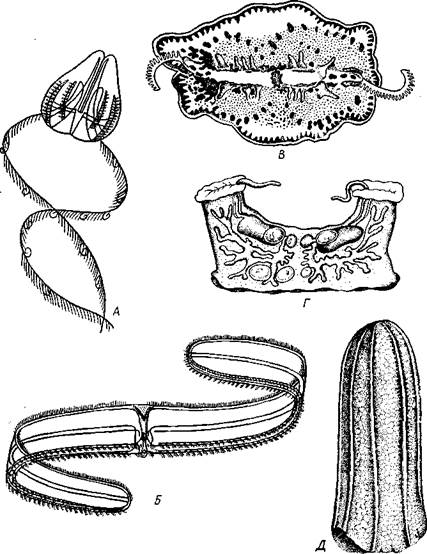
Рис. 122. Различные представители гребневиков. А — Г — щупальцевые гребневики: А — Cydippe (по Хуну): Б — «Венерин пояс» (Cestus veneris) (по Мейеру); В — ползающий гребневик Coeloplana (по Кюкенталю); Г — сидячий гребневик Tjalfiella (из Догеля); Д — бесщупальцевый гребневик Beroe cucumis (по Наумову)
ресничек. Развитие гребневиков обнаруживает следы закладки мезодермы— этим они приближаются к червям. Возможно, что гребневики отделились от общего с Coelenterata ствола развития до того, как последние стали сидячими животными,
РАЗДЕЛ БИЛАТЕРАЛЬНЫЕ (BILATERIA)
ТИП ПЛОСКИЕ ЧЕРВИ (PLATHELMINTHES, ИЛИ PLATODES)
1. Тип плоских червей представлен двусторонне-симметричными (билатеральными) животными, через тело которых можно провести только одну плоскость симметрии. Двусторонняя симметрия впервые появляется именно в этой группе беспозвоночных.
Плоские черви трехслойны. В процессе онтогенеза у них формируются не два, как у кишечнополостных, а три зародышевых листка. Между эктодермой, образующей покровы, и энтодермой, из которой построен кишечник, у них имеется еще и промежуточный зародышевый листок — мезодерма. Тело их в большинстве случаев вытянуто в длину и сплющено в спинно-брюшном направлении (принимает вид листа, пластинки, ленты).
2. Важная особенность строения плоских червей — наличие у них кожно-мускульного мешка. Так называется совокупность эпителия и расположенной непосредственно под ним сложной системы мышечных волокон. Эти волокна, нередко распадающиеся на несколько слоев (кольцевые, продольные), одевают под эпителием все тело животного в виде сплошного мешка, а не разбиваются на отдельные мускульные пучки более специального назначения, как у высших билатеральных животных (членистоногих, моллюсков). Сокращением мышечных элементов кожно-мускульного мешка обусловливаются характерные «червеобразные» движения Plathelminthes.
3. Тело плоских червей не имеет полости — это бесполостные, или паренхиматозные, животные: пространство между внутренними органами заполнено соединительной тканью мезодермального происхождения, или паренхимой, содержащей многочисленные клетки. Паренхима занимает все промежутки между органами, и ее роль многообразна. Она имеет опорное значение, служит местом накопления запасных питательных веществ, играет важную роль в процессах обмена и т. д.
4. Пищеварительный канал имеет еще примитивное устройство, состоя лишь из эктодермальной передней кишки, или глотки, и энтодер-мальной средней кишки, замкнутой слепо. Задней кишки и заднепроходного отверстия нет. У паразитических форм пищеварительная система может полностью редуцироваться.
5. Нервная система состоит из парного мозгового ганглия и идущих от него кзади нервных стволов, соединенных кольцевыми перемычками. Особого развития достигают два продольных ствола (боковые или брюшные). У плоских червей формируется центральный регулирующий аппарат нервной системы.
6. Кровеносная и дыхательная системы отсутствуют.
7. Впервые появляются специальные органы выделения, построенные по типу так называемых протонефридиев. Они представлены системой
разветвленных канальцев, оканчивающихся в паренхиме особой звездчатой клеткой с пучком ресничек. С внешней средой протонефридии сообщаются при помощи специальных экскреторных (выделительных) отверстий.
8. Половая система плоских червей гермафродитна; как правило, формируется сложная система протоков, служащих для выведения половых продуктов, и появляются органы, обеспечивающие возможность внутреннего оплодотворения.
К типу плоских червей принадлежат 5 классов: ресничные черви (Turbellaria), сосальщики, или трематоды (Trematoda), моногенеи (Мо-nogenoidea), ленточные черви (Cestoda) и цестодообразные (Cestoda-ria). Последние 4 класса представлены исключительно паразитическими формами.
КЛАСС I. РЕСНИЧНЫЕ ЧЕРВИ (TURBELLARIA)
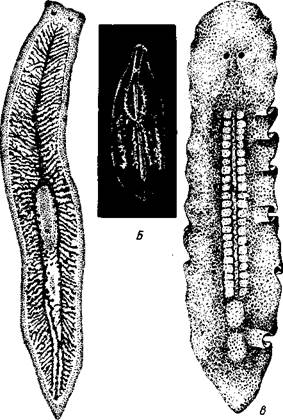
Свободноживущие морские или пресноводные, редко наземные, черви, все тело которых покрыто мерцательным эпителием (рис. 123). Число видов турбеллярий достигает 3000.
|
Строение и физиология.Тело вытянуто в длину, листовидно и обыкновенно лишено каких бы то ни было придатков. Лишь у немногих турбеллярий на переднем конце тела имеются небольшие щупальцевидные выросты (рис. 124). Величина турбеллярий незначительна и варьирует от миллиметра до нескольких сантиметров. Тело лишь в редких случаях бесцветно или сплошного белого цвета; обычно животное окрашено в разные, иногда очень яркие цвета зернами залегающего в коже пигмента.
| Рис. 123. Ресничные черви (Turbellaria). A — Dendrocoelum lacteum (Tricladida); Б — Mesosto- та ehrenbergi (Rhabdocoeia); В — Leptoplana al- cinoi (Polycladida) (по Стрелкову) |
Тело покрыто однослойным мерцательным эпителием (рис. 125, А), который имеет вид высоких цилиндрических клеток, несущих по свободному краю реснички. У некоторых турбеллярий границы между отдельными клетками исчезают. В таких случаях участки цитоплазмы, несущей ядра, иногда довольно глубоко
погружаются в толщу тканей, приобретая вид
колбочек, которые суженной частью соединены с наружным слоем (рис. 125,5). Последний лишен ядер и представляет собой неразделенную цитоплазматическую пластинку с ресничками по свободному краю. Такой тип строения покровов называется погруженным эпителием.
|
| 16 |
| 15 |
С покровами турбеллярий связаны многочисленные кожные железы. Обычно это бокаловидной или грушевидной формы железистые клетки, разбросанные по всей поверхности тела или собранные в комплексы. Они могут залегать как в самом эпителии, так и погружаться в глубь тела. Одной из разновидностей кожных желез являются рабдитные клетки, содержащие блестящие, сильно преломляющие свет палочки — раб-диты. Последние лежат перпендикулярно к поверхности тела. При раздражении животного они легко выбрасываются из эпителия наружу и, сильно разбухая в воде, образуют на поверхности тела червя сгустки слизи, возможно, играющей защитную роль. Под эпителием находится эластичная основная, или базальная, мембрана — продукт выделения оснований эпителиальных клеток; она служит для придания телу определенной формы и для прикрепления мышц. Под базальной мембраной помещается сложная мышечная система, состоящая из нескольких слоев гладких мышечных волокон. Наиболее поверхностно располагается слой кольцевых мышц, несколько глубже — продольных и, наконец, диагональных мышечных волокон, идущих в двух направлениях и перекрещивающихся между собой под углом. Совокупность мышц вместе с эпителием образует кожно-мускульный мешок. Кроме того, у всех турбеллярий имеются характерные для них дорзовентральные, или спинно-брюшные, мышцы. Это пучки волокон, идущих от спинной стороны тела к брюшной и прикрепляющихся к базальной мембране обеих плоских сторон тела; таким образом, животное может сплющивать тело в спинно-брюшном направлении.
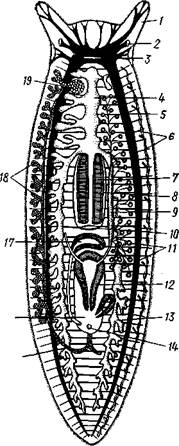
| Рис. 124. Схема строения трехвет-вистой" турбеллярий (по Граффу): 1 — щупальцевидные выросты, 2—глаза, 3 — мозговой ганглий, 4 — передняя ветвь кишечника, 5 — продольный нервный ствол, 6 — поперечные нервные перемычки, 7 — глотка, S — глоточный карман, 9 — семяпровод, 10—ротовое отверстие, // — семенники, 12 — совокупительный орган, 13 — задняя ветвь кишечника, 14 — половое отверстие, 15 — яйцевод, 16—половая клоака, 17 — комулятивная сумка, 18 — желточ-ники, 19 — яичник. Слева удалены семенники, справа — желточники и яичник |
Движение турбеллярий, с одной стороны,— результат биения ресничек наружного эпителия, с другой — обусловливаются сокращением кожно-мускульного мешка. Turbellaria и ползают, и плавают. Мелкие формы главным образом плавают при помощи ресничек. Более крупные представители ползают, вытягивая, сокращая и изгибая тело.
Кнутри от кожно-мускульного мешка все промежутки между органами заполнены паренхимой — рыхлым скоплением соединительнотканых клеток, соединяющихся друг с другом своими отростками.
Между клетками имеются небольшие пространства с водянистой жидкостью, что позволяет паренхиме выполнять роль посредника в передаче продуктов пищеварения от кишечника к внутренним органам. Одновременно осуществляется и перенос продуктов обмена к выделительной системе. Кроме того, наличие многочисленных фибриллярных образований позволяет рассматривать паренхиму как опорную ткань.
Пищеварительная система состоит из передней и средней кишки, замыкающейся слепо. Рот служит не только для проглатывания пищи,
|
|
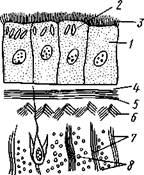
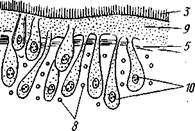
Рис. 125. Схема строения кожно-мускульного мешка. А —с типичным
мерцательным эпителием, Б — с погруженным эпителием (по Граф-
фу, схематизировано):
Клетки эпителия, 2 — рабдиты, 3 — реснички, 4 — базальная мембрана, 5 — кольцевые мышцы, 6 — диагональные мышцы, 7 — дорзовентральные мышцы. 8 — продольные мышцы, 9 — наружный цитоплазматический слой, 10 — погруженные участки цитоплазмы с ядрами
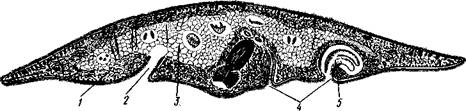
Рис. 126. Строение бескишечной турбеллярии Convoluta (по Бреслау):
Дата добавления: 2015-01-24; просмотров: 1722;