Роль связывания в плазме
Циркулируя в крови, половые и надпочечниковые стероиды присутствуют в виде свободных гормонов и связанных форм, образующих комплексы с белками плазмы. Хотя секретируемые стероиды в тех низких концентрациях, которые содержатся в плазме (примерно 10~9 M), растворимы в воде, основная масса циркулирующих гормонов транспортируется в виде стероидбелковых комплексов [71]. С белками плазмы связано более 98% половых стероидов, около 90% содержащегося в крови кортизола и около 50% альдостерона. Поскольку на уровне клеток-мишеней активны именно свободные гормоны, роль связывания в плазме может заключаться в создании резервуара или буфера, который контролирует доступность гормона для рецепторов клеток-мишеней. Хотя каждый стероидный гормон может связываться тем или другим белком плазмы, но степень и прочность (или сродство) связывания разных стероидов значительно варьируют. Так, около 40% содержащегося в крови альдостерона слабо связано с сывороточным альбумином, тогда как более 90% кортизола и тестостерона связаны с высоким сродством специфическими белками, называемыми соответственно кортикостероидсвязывающим глобулином (КССГ), или транс-
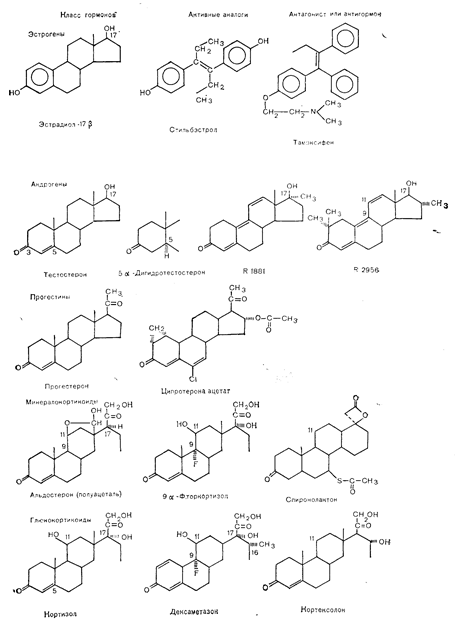
Рис4—25. Структура главных классов стероидных гормонов. Лроизводные андрогенов R 1881 и R 2956 носят кодовые номера фармацевтической компании Roussel (Mainwaring [70] в модификации). кортином, и сексгормонсвязывающим глобулином. Последний белок связывает и эстрадиол, хотя в физиологических условиях, вероятно, степень его связывания не слишком велика.
В отношении типичных стероидных гормонов, равно как и в отношении аналогичных гормонов, таких, как витамин D и тироксин, отсутствуют указания на важность роли их связывания в плазме в механизме биологического действия. Как уже отмечалось, именно свободный гормон представляет собой биологически активную фракцию и именно его уровень коррелирует с величиной реакции на гормон. Кроме того, стероиды активны и в отсутствие своих связывающих белков, а синтетические аналоги стероидов, которые обычно не связываются с транспортными белками, сохраняют способность вызывать биологические эффекты в тканях-мишенях за счет взаимодействия со специфическими внутриклеточными рецепторами. Когда в условиях патологии или под влиянием терапевтических мероприятий концентрация связывающих белков повышается или снижается, концентрация свободного стероида остается относительно постоянной, несмотря на изменение общего уровня гормона в крови.
Хотя связывание в плазме, по-видимому, не имеет непосредственного значения для гормонального эффекта (кроме значения в качестве резервуара циркулирующего стероида), оно может, очевидно, оказывать влияние на скорость клиренса крови от секретируемого, инъецируемого или принимаемого внутрь гормона. Связывание с белками плазмы может также служить буфером, сводящим к минимуму влияние эпизодической секреции гормона на концентрацию свободного стероида, поступающего к клетке-мишени. При лечении стероидными гормонами связывание с транспортными белками плазмы могло бы снижать скорость метаболизма вводимого стероида и создавать резервуар, из которого постепенно освобождается свободный гормон. Степень, в которой эти эффекты связывания в плазме влияют на продолжительность и интенсивность реакций на отдельные стероиды, не ясна, но она могла бы иметь очевидное значение для фармакокинетики стероидных препаратов [69]. Связывание с альбумином не снижает доступности стероидных гормонов для головного мозга, тогда как гормон, связанный с глобулинами, не транспортируется в мозг. Таким образом, доля содержащегося в плазме гормона, которая может транспортироваться в головной мозг, не ограничивается свободной фракцией, а включает и более крупные, связанные с альбумином, комплексы [72]. В крысиной плазме эта доля могла бы составлять 60% прогестерона, 40% тестостерона и 15% кортикостероидов, тогда как свободные фракции этих гормонов в плазме колеблются в пределах 2—8%.
Другой физиологически важный аспект связывания стероидов с белками плазмы заключается в значении избирательности этого процесса для действия альдостерона на его специфические минералокортикоидные рецепторы в почках. Поскольку главные участки связывания альдостерона в почках обнаруживают также высо- кое сродство к дезоксикортикостерону (ДОК) и меньшее, но отчетливое, сродство к кортизолу, эти стероиды могли бы, очевидно, занимать почечные рецепторы, так как их концентрации в крови близки (ДОК) или на несколько порядков выше (кортизол), чем концентрация альдостерона. Однако более 90% ДОК и кортизола, присутствующих в крови, связаны с альбумином и КССГ, тогда как только 40% альдостерона в циркуляции связано с альбумином. В связи с этим ДОК и глюкокортикоиды конкурируют за рецепторы альдостерона in vivo много слабее, чем in vitro в условиях отсутствия плазмы (например, для ДОК 1—5% против 80%). Без избирательности связывания ДОК и глюкокортикоидов в плазме альдостерон не мог бы насыщать минералокортикоидные рецепторы и оказывать характерное регулирующее действие на гомеостаз натрия. Этим способом связывание в плазме способствует предотвращению связывания «неадекватных» стероидов (таких, как ДОК, кортикостерон и кортизол, которые могут действовать как минералокортикоиды, но секретируются под контролем АКТГ) с обладающими высоким сродством л низкой емкостью рецепторными участками в клетках-мишенях минералокортикоидов [73].
Следует отметить, что циркулирующие в крови стероидсвязывающие белки в некоторых отношениях отличаются от внутриклеточных рецепторов стероидов, опосредующих реакцию клетки-мишени. С физической точки зрения, связывающие белки плазмы — это относительно стабильные, растворимые глобулярные гликопротеины, тогда как рецепторы асимметричны, сравнительно более гидрофобны и менее стабильны. С функциональной же стороны, транспортные белки связывают преимущественно природные стероидные молекулы и хуже (или вообще не связывают) более активные синтетические аналоги. Рецепторные же молекулы, напротив, связывают как природные, так и синтетические стероиды (или их метаболиты) соответственно их биологической активности.
Дата добавления: 2015-01-19; просмотров: 1169;