СЕКРЕТОРНЫЙ ПРОЦЕСС В СИМПАТИЧЕСКИХ НЕРВАХ
Высвобождение норадреналина из симпатических нервов происходит путем экзоцитоза [33, 34]. Этот процесс несколько отличается от аналогичного процесса, свойственного секреторным клеткам, продуцирующим полипептидные и белковые гормоны, тем, что из аксональной области нервной клетки выделяется не содержимое гранул, что наблюдается в клетках, продуцирующих белковый эндокринный секрет, а целиком вся окруженная мембраной гранула с ее содержимым [35]. Такой «тотальный» экзоцитоз предполагает необходимость синтеза нового фермента и компонентов мембраны для возмещения их количества, теряемого в результате секреции. Таким образом, последний этап синтеза норадреналина — -гидроксилирование — решительно зависит от синтеза белка. Ингибиторы белкового синтеза вызывают резкое прекращение продукции и секреции норадреналина. Вторым следствием такого секреторного процесса является попадание в кровь b-гидроксилазы. В настоящее время получена антисыворотка к изолированному и очищенному ферменту, и радиоиммунологический анализ с использованием этой антисыворотки представляет собой чувствительный метод определения нейросекреторной активности в условиях, когда уровень катехоламинов в крови оказывается ниже возможностей его измерения [35].
Экзоцитоз, по-видимому, находится под непосредственным контролем со стороны нервной системы. Секреторные гранулы с содержащимися в них катехоламинами выделяются в кровь в ответ на электрическое раздражение. При этом обнаруживается связь с присутствием ацетилхолина и/или входом в клетку ионов кальция и зависимость от энергии, т. е. интактности окислительных процессов или гликолиза. На миграцию секреторных гранул к периферии клетки может влиять сокращение микрофибрилл или микротрубочек. Винбластин и колхицин, известные ингибиторы функций микротрубочек, угнетают выделение катехоламинов. Не вся секреция катехоламинов происходит путем экзоцитоза»
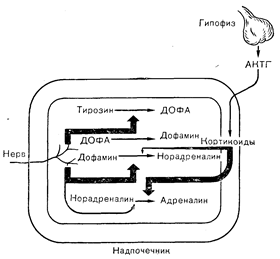
Рис. 3—15. Синтез и реууляция продукции катехоламинов в мозговом слое надпочечников. Образование ДОФА из тирозина под действием тирозингидроксилазы и Превращение дофамина в норадреналин под действием дофамина-р-гидроксилазы регулируется главным образом нервными импульсами. Метилирование норадреналина с образованием адреналина под действием фенилэтаноламин-N-Meтилтрансферазы контролируется в основном дгортикоидами (Axelrod J., Weinshilboum R., N. Engl. J. Med., 1972, 287, 237).
Небольшие их количества непрерывно «просачиваются» в кровь, причем в некоторых условиях необычные вещества, такие, как ацетальдегид, которые накапливаются в организме в результате метаболизма алкоголя, могут способствовать выделению катехоламинов без экзоцитоза. Симпатомиметические амины, такие, как тирамин и амфетамин, стимулируют секрецию катехоламинов и тем самым вызывают физиологически важные прессорные эффекты, что сопровождается истощением тканевых запасов катехоламинов. Резерпин также истощает тканевые запасы катехоламинов, но механизм его действия отличается от механизма действия симпатомиметических аминов. Резерпин ингибирует связывание норадреналина с мембранами гранул, и освобожденный норадреналин диффундирует из гранул и метаболизируется в цитоплазме под действием внутринейрональной моноаминоксидазы. Выделяемые в кровь метаболиты биологически инертны и не вызывают периферических прессорных реакций.
Дата добавления: 2015-01-19; просмотров: 1296;