СТРОЕНИЕ И ФУНКЦИИ МЕХАНИЧЕСКОЙ ТКАНИ
Функцию «внешнего скелета» у крупных древесных растений в некоторой степени выполняют перидерма и корка, «одевающие» растение снаружи. Но только механические ткани, появившиеся у растений в результате приспособления к наземной жизни, взаимодействуя с другими тканями организма, образуют прочный «внутренний скелет» растения, его арматуру. Степень развития механических тканей во многом зависит от условий обитания: они почти отсутствуют у растений влажных лесов, у многих прибрежных растений, но зато хорошо развиты у большинства растений засушливых местообитаиий. Все механические ткани можно разделить на три группы, в зависимости от строения и выполняемых функций: колленхима, склеренхима и склереиды.
КОЛЛЕНХИМА - паренхимная ткань, появляется только как первичная ткань и служит для укрепления молодых стеблей и листьев, когда продолжается растяжение клеток в длину. Обеспечивая прочность органу, колленхима в то же время способна растягиваться по мере роста молодых органов. Пластичное растяжение оболочек возможно лишь при активном воздействии живого содержимого, которое выделяют вещества, снижающие упругость оболочек. Оболочка клеток колленхимы характеризуется высоким содержанием целлюлозы (около 30 %) и гемицеллюлоз (свыше 50 %), воды, бедна пектиновыми веществами, лигнин в ней обычно отсутствует. Особенность оболочки - многослойность. Другая особенность клеток состоит в том, что выполнять свои функции арматурной ткани они могут только в состоянии тургора. При потере молодыми органами воды тонкие участки оболочек складываются гармошкой, листья и стебли теряют упругость и обвисают, т. е. завядают.
Располагается колленхима почти всегда по периферии органа или сплошным слоем или собрана отдельными пучками, связанными с проводящей тканью.
Клетки колленхимы характеризуются неравномерным утолщением стенок, за счет чего они могут выполнять опорную функцию. В оболочке наблюдаются редкие простые поры, пронизанные многочисленными плазмодесмами.
Характерное утолщение стенок клеток колленхимы носит название «утолщение колленхиматозного типа». В зависимости от характера утолщений стенок и соединения клеток между собой, различают уголковую, пластинчатую и рыхлую колленхиму.
В уголковой (наиболее распространенной) оболочка сильно утолщается в углах, где сходятся соседние три-пять клеток. Утолщенные части оболочек обычно сливаются между собой, так что границы отдельных клеток обнаруживаются с трудом.
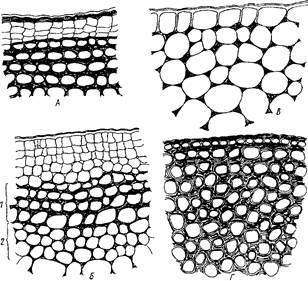
Рис. Типы колленхимы: А - пластинчатая колленхима в однолетнем побеге черешчатого дуба - Quercus longipes под формирующейся пробковой тканью, Б - пластинчатая (1) и уголковая (2) колленхима в стебле баклажана - Salanum melongena, В - уголковая колленхима в черешке листа бегонии - Begonia rex, Г — рыхлая колленхима в черешке листа лопуха большого - Arctium platylepis
В пластинчатой колленхиме сплошными параллельными слоями утолщаются тангенциальные стенки, радиальные же остаются тонкими. Чаще всего эти слои параллельны поверхности органа.
В рыхлой колленхиме утолщение оболочек происходит на тех участках стенок, которые примыкают к межклетникам.
Колленхима широко распространена у двудольных, у однодольных если и встречается, то в области стеблевых узлов соломины злаков, что предохраняет растения от полегания. Колленхима характерна для растущих травянистых стеблей, черешков листьев, цветоножек и т. д. Обычно она располагается в несколько слоев по периферии органа под эпидермисом. У древесных пород колленхиму можно наблюдать до разрушения первичной коры, перед появлением перидермы. Особую форму колленхимы представляет гиподерма листа (субэпидермальный слой клеток), выполняющая наряду с механической функцией функцию дополнительной защиты листа от излишнего испарения (например в хвое сосны или ели).
СКЛЕРЕНХИМА - наиболее важная механическая ткань, которая встречается в органах почти всех высших растений. Клетки ее имеют прозенхимную форму и представляют собой волокна, длинные, плотно расположенные, с заостренными концами и равномерно утолщенной, чаще всего одревесневшей оболочкой. Живое содержимое клеток обычно отмирает. Отдельные клетки склеренхимы называются элементарными волокнами. Отдельный тяж элементарных волокон (склеренхимных волокон) называют техническим волокном. Волокна встречаются повсеместно во всех органах в виде отдельных тяжей (пучков) или сплошным цилиндрическим кольцом, группами или рассеянно в проводящей ткани.
По сравнению с колленхимой склеренхимные волокна отличаются большей упругостью, равной 15-20 кг/мм2, тогда как у колленхимы она составляет не более 10-12 кг/мм2. Наличие склеренхимы дает возможность органам растения противостоять нагрузкам, которые возникают в результате изгиба или под действием массы самого растения.
По происхождению различают первичную и вторичную склеренхиму. Первичная дифференцируется из клеток основной меристемы апексов, прокамбиальных пучков или из перициклических волокон. Вторичную формирует камбий. В зависимости от расположения в теле растения и функциональных особенностей волокна подразделяют на две группы: древесинные (ксилемные) волокна, входящие в состав проводящей ткани ксилемы, и лубяные (экстраксилярные) волокна различных систем тканей вне ксилемы.
Древесинные волокна, или либриформ, не более 2 мм, одревесневшие оболочки снабжены простыми порами со щелевидными очертаниями, Либриформ называют многофункциональной тканью, что связано с изменчивостью морфоструктуры волокон. Встречаются переходные элементы между клетками либриформа и водопроводящими элементами. В этом случае клетки либриформа принимают участие в транспортировке воды. У таких волокон рано отмирает живое содержимое. Наблюдаются переходные формы между клетками либриформа и древесинной паренхимой. В этом случае клетки волокон сохраняют живое содержимое, в таких клетках запасаются крахмал и другие органические вещества.
Клетки либриформа очень прочны, но почти неэластичны. Главная его функция - опора для водопроводящих элементов и для всего растения. Этот тип склеренхимы широко распространен среди высших растений. У лиственных деревьев либриформ занимает значительную часть древесины, особенно в тех ее массивах, которые сформировались во вторую половину вегетационного периода.
Лубяные волокна расположены обычно в коровой части осевого органа, во флоэме, они встречаются в коре стебля и корня, а также в листовых черешках и пластинках, в цветоножках, плодоножках. Клетки лубяных волокон длинные, толстостенные. Длина их колеблется от 40 до 60 мм, у китайской крапивы рами от 350 до 500 мм (субтропическое прядильное растение). В целом волокна насыщены цитоплазмой, в ней содержатся единичные мелкие хлоропласты, часто с крахмальными зернами. Число митохондрий значительно больше, чем хлоропластов. Характерная черта молодых волокон - высокоактивный комплекс Гольджи. У сформировавшихся лубяных волокон протопласт чаще всего отмирает и полость клетки совершенно исчезает.
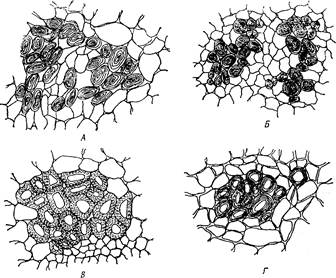
Рис. 8. Лубяные волокна у различных растений (поперечный срез):
А - в стебле конопли посевной - Cannabis sativa, Б - в побеге щелковицы, белой - Morus alba. В—в черешке листа тополя канадского — Populus canadensis, Г — в стебле льна обыкновенного - Linum usitatissimum
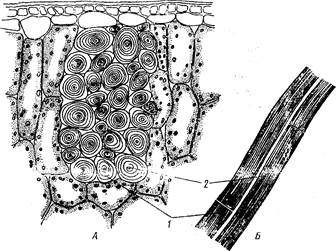 |
Рис. 9. Лубяные волокна в листьях пальмы веерной -Trachycarpus excelsa:
A - поперечный, Б - продольный срезы: 1 — оболочка, волокна, 2 — полость волокна
За счет интрузивного роста лубяных волокон создается исключительная прочность ткани, которая повышается благодаря спиральным расположениям микрофибрилл.
Свойства лубяных волокон (прочность, исключительная эластичность, большая длина волокна, отсутствие одревеснения) очень ценны для текстильной промышленности. Особый интерес представляют такие растения, как лен, конопля, кенаф, джут, рами, их техническое волокно используется в текстильной промышленности.
Текстильные качества лубяных волокон неодинаковы. Различают твердо- и мягковолокнистые растения. У мягковолокнистых (лен) волокна мягкие, эластичные, идут на изготовление высококачественных льняных тканей. У твердоволокнистых (новозеландский лен, виды агавы, некоторые пальмы) волокна твердые, грубые, из них делают веревки, щетки, грубые волокнистые ткани.
СКЛЕРЕИДЫ представлены клетками самой разнообразной формы, с равномерно утолщенными слоистыми стенками, пронизанными порами. Стенки склереид всегда сильно одревесневают, иногда пропитываются известью, кремнеземом и кутином. Живое содержимое, как правило, отмирает.
Склереиды встречаются в различных органах растений: плодах, листьях, стеблях, могут формироваться на протяжении всего онтогенеза. Группы склереид бывают рассеяны в мякоти плодов, либо частично перемешаны с паренхимными клетками, либо составляют плотную, без межклетников, ткань (косточки плодов сливы, черешни, абрикоса, скорлупа ореха и т. д.).
Классификация склереид основана на их морфоструктуре. Различают следующие типы: брахисклереиды, или каменистые клетки, -короткие изодиаметрические склереиды, широко распространены в коре, лубе и сердцевине стеблей, а также мякоти плодов; макросклереиды - удлиненные палочкообразные склереиды, есть в семенах бобовых; остеосклереиды - напоминают трубчатую кость, присутствуют в листьях многих двудольных и в семенной кожуре; астросклереиды (звездчатые склереиды) - разветвленные клетки, находятся в листьях двудольных; трихосклереиды - ветвистые тонкостенные склереиды, напоминающие волоски растений, ответвления которых проникают в межклетные пространства. Особую форму одиночных склереид, встречающихся в кожистых и мясистых листьях, представляют опорные клетки - идиобласты, они несут всю тяжесть сочной хлоренхимы листьев и часто встречаются в листьях вечнозеленых растений (чай, камелия, маслины), а также у облепихи.
В большинстве случаев склереиды возникают непосредственно из клеток апикальной меристемы стебля либо из клеток, лежащих под наружным слоем меристематических клеток, превращающихся в эпидермис. Они могут формироваться из обычных паренхимных клеток первичной коры или сердцевинных лучей путем утолщения и последующего одревеснения их стенок.
 Склереиды не всегда играют чисто механическую функцию. Так, например, в коре деревьев и кустарников они укрепляют склеренхим-ную арматуру и вместе с тем защищают кору от поедания травоядными животными.
Склереиды не всегда играют чисто механическую функцию. Так, например, в коре деревьев и кустарников они укрепляют склеренхим-ную арматуру и вместе с тем защищают кору от поедания травоядными животными.
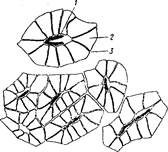
Рис. 10. Склереиды по углам складок листовых бугорков на конусах нарастания стебля дуба длинноножкового:
1 — полость склереида, 2 - оболочка, 3 — пора.
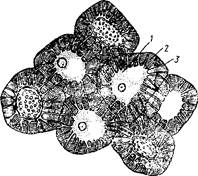
Рис. 11. Склереиды косточки неспелой алычи растопыренной с живым содержимым:
1 - цитоплазма, 2 - утолщенная слоистая оболочка,
3 - ветвистая пора на стенке склереида
ПРОВОДЯЩИЕ ТКАНИ И КЛАССИФИКАЦИЯ ПРОВОДЯЩИХ ПУЧКОВ
При выходе растений на сушу наряду с проблемой прочности организма возникла проблема снабжения органов и тканей питательными веществами. Вследствие этого появились специализированные группы клеток для проведения необходимых растению веществ - проводящие ткани. Наземное растение получает питание из почвы (почвенное питание) и из окружающей атмосферы (воздушное питание). Для обеспечения транспорта этих токов возникли две проводящие ткани, по которым вещества передвигаются в двух противоположных направлениях. Ксилема проводит вещества почвенного питания, воду с растворенными минеральными солями в направлении снизу вверх, т. е. осуществляет восходящий ток. Флоэма проводит вещества, синтезируемые листьями, в направлении сверху вниз, т.е. осуществляет нисходящий ток. Являясь узкоспециализированными тканями, ксилем и флоэма имеют ряд общих признаков:
1. Представлены в теле растения непрерывной системой, разветвленной и соединяющей все органы растения — от самых молодых побегов до тончайших корневых волосков.
2. Относятся к сложным тканям, т. к. в их состав входят морфологически и функционально разнородные элементы - проводящие, запасающие, механические и выделительные. Каждый из этих элементов выполняет определенную функцию, но все вместе они обусловливают проведение веществ. Основными являются проводящие элементы, остальные обеспечивают выполнение их функций.
3. Проводящие комплексы флоэмы и ксилемы представлены прозенхимными, вытянутыми вдоль осевых органов высокоспециализированными элементами, стенки которых содержат поры или перфорации, облегчающие прохождение веществ.
4. По происхождению и развитию проводящие ткани могут быть первичными и вторичными. Первичные проводящие ткани дифференцируются в процессе развития первичного тела растения меристема-тической тканью прокамбием, вторичные образуются камбием и связаны с важной стадией развития растения - увеличением толщины осевых органов.
5. В большинстве случаев ксилема и флоэма располагаются рядом, образуя проводящие пучки.
КСИЛЕМА (древесина) - основная водопроводящая ткань сосудистых растений. Ксилема состоит из нескольких типов клеток. Основными являются трахеиды и трахеи (членики сосуда), вспомогательными - клетки механической ткани и клетки паренхимы.
Трахеиды — прозенхимные клетки длиной в несколько миллиметров, шириной в десятые и сотые доли миллиметра. Взрослые трахеиды представляют собой мертвые клетки с утолщенными одревесневшими оболочками, несущими поры, часто окаймленные. Фильтрация растворов из одной трахеиды в другую происходит через эти поры. Формируются трахеиды из прокамбиальных пучков верхушечной меристемы, а также из камбия. Для них характерен интрузивный (скользящий) рост, когда одна из трахеид внедряется своим заостренным концом между другими трахеидами. Образуется сплошная масса плотно прилегающих друг к другу трахеид, заполняющих большую часть объема осевых органов растений. Трахеиды передают растворы не только в вертикальном, но и в горизонтальном направлениях, в лежащие рядом проводящие и паренхимные клетки. Поэтому боковые стенки у них водопроницаемы. В то же время для трахеид характерны утолщения различной формы - кольчатые, спиральные, сетчатые, лестничные и точечные. Такое утолщение оболочки, не по всей ее поверхности, а в отдельных участках, имеет огромное биологическое значение. При относительно экономном расходовании органического вещества водопроводящие элементы оказываются устойчивы к сжатию и растяжению и в то же время проницаемы (в поперечном направлении).
У папоротников и голосеменных трахеиды служат единственным проводящим элементом в ксилеме. У покрытосеменных трахеиды перемежаются с сосудами и другими элементами ксилемы. У многих покрытосеменных трахеиды вообще отсутствуют, т. к. их наличие считается признаком примитивности и древности вида.
Кроме проводящей, трахеиды выполняют механическую функцию. У голосеменных и некоторых цветковых растений специальные механические элементы отсутствуют, прочность стебля обеспечивается трахеидами. В стволах хвойных, например, один или два наружных слоя трахеид выполняют функцию транспортировки воды, все внутренние слои, т. е. почти вся масса трахеид, воду не проводят и функционируют как механическая ткань. Между трахеидами и механическими волокнами (либриформом) существует ряд переходных форм.
Сосуды, или трахеи, состоят из многих клеток, которые называются члениками сосуда. Членики располагаются друг над другом, образуя длинную полую трубку. Поперечные перегородки между члениками растворяются, и возникают перфорации (сквозные отверстия). По таким полым трубкам растворы передвигаются значительно легче, чем по трахеидам. Каждый сосуд может состоять из огромного числа члеников, поэтому средняя длина сосудов - несколько сантиметров (иногда до 1 метра и более). Пример: у дуба длина сосуда достигает 2 м, лиан - 3—5 м, иногда несколько десятков метров.
Типы члеников сосуда различны. Самые совершенные состоят из широких коротких члеников, диаметр которых превышает длину, а в перфорационных пластинках имеется одно крупное отверстие (дуб). Менее специализированные сосуды состоят из более длинных и узких члеников, поперечные стенки между которыми наклонны. Перфорационные пластинки имеют несколько отверстий, расположенных друг над другом (лестничная перфорация). Существует известная корреляция между признаками: чем длиннее членики сосуда, тем они уже, тем более наклонна конечная стенка, тем мельче и многочисленнее перфорации.
По характеру утолщений клеточных стенок различают кольчатые, спиральные, лестничные, сетчатые и поровые сосуды.
Кольчатые и спиральные сосуды в процессе онтогенеза растений появляются первыми и функционируют относительно недолго. Эти сосуды мелкие и тонкостенные, стенки их одревесневают лишь частично, они могут растягиваться и поэтому не препятствуют росту органа. Такой тип сосудов у всех покрытосеменных появляется раньше других и вместе с трахеидами образует проводящие элементы протоксилемы. Уже в метаксилеме, а тем более во вторичной ксилеме ни кольчатые, ни спиральные сосуды не встречаются.
Сетчатые и точечные сосуды более крупные и более прочные, появляются в онтогенезе позднее, во вторичной ксилеме, и функционируют в течение года и более.
Древесинные волокна (либриформ) - это мертвые вытянутые клетки с одревесневшими оболочками, создающие опору и защиту водопроводящим элементам и клеткам паренхимы. Эволюционно возникли, как и сосуды, из трахеид. Но преобразование трахеид шло в ином направлении: водопроводящая функция постепенно угасала, окаймленные поры превратились в небольшие по размерам простые поры, повысившие механическую прочность оболочек.
Паренхимные элементы - живые клетки, составляющие иногда до 25 % и более общего объема древесины. Клетки паренхимы являются причастными в той или иной степени к транспорту по ксилеме. Паренхимные клетки часто окружают сосуды, образуя обкладку (контактная паренхима). Околососудистые клетки находятся в тесном контакте с растворами, заполняющими сосуды, и, в зависимости от условий, то конденсируют в своих пластидах поступающие к ним углеводы и другие вещества, то отдают их в полость сосуда. Осенью и перед распусканием листьев в таких клетках находится в изобилии крахмал, количество которого в другие периоды значительно уменьшается. Сообщаются паренхимные клетки обкладки с сосудами через односторонние окаймленные поры. Таким образом, ксилема - это единая система, в которой сосуд - элемент проведения растворов, околососудистые клетки паренхимы регулируют поступление растворов, направление и скорость их движения, а либриформ придает прочность этой системе. Наиболее специализированными клетками паренхимы являются передаточные клетки, обладающие своеобразными выростами клеточных стенок (протуберанцы). Эти клетки сопровождают трахеальные элементы в местах наиболее интенсивного выхода раствора в окружающие ткани.
В корне околососудистые паренхимные клетки иногда склерифицируются, и в связи с этим в процессе развития обнаруживаются общие черты с трахеальными элементами. Кроме опорной функции, они могут выполнять транспорт водных растворов в радиальном направлении. Собранные в горизонтальные полосы участки паренхимы образуют так называемые сердцевинные или ксилемные лучи.
Клетки паренхимы, примыкающие к сосуду, могут образовывать выросты в полость сосудов через поры - тилы (греч. - вздутие, утолщение). Со временем они разрастаются, заполняют всю полость сосуда. Ядро паренхимной клетки либо переходит в тилу, либо делится, при этом одно ядро остается в паренхимной клетке, другое переходит в тилу. Дальнейшее деление ядра в тиле приводит к образованию ложной паренхимной ткани, закупоривающей полость сосуда. Этот процесс называется тидо-образованием, он характерен в основном для многолетних древесных растений и связан с возрастными изменениями древесины. В тилах откладываются запасные пластические вещества, так же как в клетках древесинной паренхимы, иногда - конечные продукты обмена веществ, вследствие чего клетка погибает. В некоторых случаях они плотно заполняют полость сосуда, оболочки их сильно утолщаются, пронизываются перовыми каналами. Тилы становятся похожими на каменистые клетки, заполненные различного рода отложениями. Такие тилы играют особую роль в процессе формирования ядра древесины.
Тилообразование имеет важное биологическое значение, т. к. усиливает механическую прочность центральной части стволов деревьев, консервирует их смолоподобными органическими соединениями, не поддающимися воздействию микробов и насекомых.
ФЛОЭМА - важнейшая проводящая пластические вещества ткань сосудистых растений. Представлена комплексом тканей: проводящей, которая включает ситовидные трубки и клетки спутницы;
основной, представленной паренхимными клетками, и механической, состоящей из лубяных волокон со склереидами.
Флоэмная ткань менее склерифицирована и менее долговечна, чем ксилема. Флоэма занимает периферическое положение в стебле и корне, поэтому она претерпевает значительные изменения при увеличении окружности осевых органов и в конечном счете сминается перидермой. Старая ксилема, наоборот, в структурном отношении остается относительно неизменной.
Ситовидные элементы наблюдаются двух типов. Более примитивный - ситовидная клетка, которая представляет собой сильно вытянутую в длину клетку, с заостренными концами, на боковых стенках содержит ситовидные поля. Ситовидные поля представляют собой участки клеточной стенки, пронизанные многочисленными отверстиями, через которые проходит сообщение протопластов соседних клеток. Другим примитивным признаком ситовидных клеток является отсутствие сопровождающих клеток. Пластические вещества проходят через боковые ситовидные поля из одной клетки в другую. Такой тип ситовидных элементов присущ высшим споровым и голосеменным растениям.
Ситовидные трубки — высокоспециализированные элементы. Каждая трубка состоит из вертикального ряда живых вытянутых клеток — члеников, соединенных между собой поперечными стенками — ситовидными пластинками (участки общих стенок члеников, несущие ситовидные поля). Ситовидные трубки обычно тянутся вдоль продольной оси органа, но есть и поперечно идущие, соединяющие группы проводящих тканей. Оболочки ситовидных трубок целлюлозные, лишь к концу вегетации некоторые трубки одревесневают. В полостях ситовидных трубок долго сохраняется живой протопласт в виде пристенного слоя, ядро в зрелых элементах отсутствует, живут клетки, как правило, одну вегетацию. Около ситовидных трубок имеются сопровождающие клетки, структурно и функционально связанные с ними.
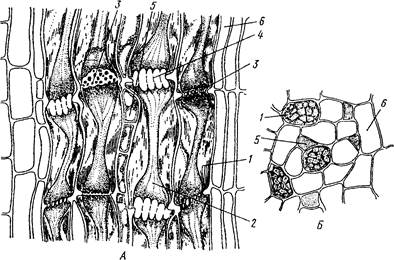
Рис. 14. Ситовидные элементы, во флоэме тыквы обыкновенной:
А — продольный, Б — поперечный срезы: 1 — функционирующая ситовидная трубка, 2 - ситовидная трубка с закупоренными ситовидными поперечными перегородками (мозолистое тело), 3 — незакупоренные ситовидные поперечные перегородки, 4 - закупоренные ситовидные поперечные перегородки, 5 — клетка-спутница, 6 — лубяная (флоэмная) паренхима
Клетки-спутницы образуются из одной материнской клетки с ситовидной трубкой. Это паренхимные элементы флоэмы, обеспечивающие регуляцию передвижения веществ по флоэме, они связаны с ситовидными трубками плазмодесмами. Протопласты клеток-спутниц отличаются метаболической активностью: ядро и ядрышко крупные, множество хлоропластов, крупных митохондрий, рибосом, имеется эндоплазматический ретикулум. Клетки вакуолизированы. Особенностью клеточных стенок является не только наличие многочисленных ветвящихся плазмодесм, но и обилие в них инвагинаций (лабиринтов), выстланных плазмалеммой, что значительно увеличивает поверхность. Активный метаболизм в клетках-спутницах, тесная связь их с ситовидными трубками, в значительной степени утративших признаки самостоятельных живых систем, указывают на активную роль сопровождающих клеток во флоэмном транспорте. Клетки-спутницы интенсивно вырабатывают различные ферменты, которые передаются в ситовидные трубки, служат также источником энергии для транспорта веществ в ситовидных трубках.
Основными веществами флоэмного тока служат сахара, главным образом сахароза, кроме того обнаружены азотистые вещества (белки, аминокислоты - не более 0,5 %), органические кислоты, витамины, фитогормоны. Передвижение пластических веществ (ассимилятов) по ситовидным трубкам происходит со скоростью 50-100 см/ч и включает в себя три взаимосвязанных процесса: загрузку флоэмы, транспорт ассимилятов и разгрузку флоэмы. Передвижение ассимилятов требует интенсивного дыхания клеток флоэмы, если дыхание затруднено, то передвижение останавливается.
Лубяная паренхима - постоянный компонент флоэмы. В клетках лубяной паренхимы активно протекают обменные реакции и накапливаются различные эргастические вещества - крахмал, жиры, различные органические соединения, а также таннины, смолы, кристаллы. Клетки паренхимы имеют продолговатую форму и располагаются параллельно ситовидным трубкам. Связь осуществляется посредством простых пор, которые собраны группами. Оболочки паренхимных клеток целлюлозные, тонкие, физиологическая активность высокая, расположение среди ситовидных элементов беспорядочное.
Вторичная флоэмная паренхима подразделяется на две системы:
вертикальную и горизонтальную. В вертикальной системе лубяная паренхима располагается вместе с ситовидными и механическими элементами, горизонтальная система представляет паренхиму сердцевинных лучей. Такая структура особенно характерна для древесных растений.
Склеренхимные элементы представлены лубяными волокнами и склереидами. Волокна относятся к обычным компонентам первичной и вторичной флоэмы. В первичной волокна встречаются только в периферической части ткани, во вторичной - распределены по всей ткани среди других клеток осевой системы. В зрелом состоянии волокна могут быть живыми и мертвыми, одревесневшими или неодревесневшими. Живые волокна выполняют функцию запаса. У многих видов первичные и вторичные лубяные волокна используются как источники промышленного волокна.
Во флоэме часто встречаются склереиды. Они располагаются в комбинации с волокнами либо отдельной группой и в осевой, и в лучевой системах вторичной флоэмы. Как правило, склереиды образуются в более старых участках флоэмы вследствие склерификации паренхим-ных клеток. Нередко склерификации предшествует интрузивный рост клеток, формируются нитчатые и разветвленные склереиды. Длинные и тонкие склереиды напоминают волокна и часто называются волокнистыми склереидами.
ПРОВОДЯЩИЕ ПУЧКИ
Проводящие элементы в комплексе с паренхимными и механическими элементами образуют в теле растения тяжи, которые называют проводящими пучками.
В молодых органах большинства растений проводящие пучки идут раздельно. На более поздних стадиях развития органов проводящие пучки сливаются, образуя сплошной цилиндр, состоящий из тканей древесины и луба. У однодольных пучковая структура сохраняется на всех стадиях развития органа.
Система проводящих пучков совершенствовалась в процессе приспособления растений к наземным условиям существования. Она пронизывает все органы растений, объединяя их в одно целое и обеспечивая в растении единый обменный процесс со множеством разнообразных отправлений.
Формирование проводящих пучков осуществляется на ранних стадиях развития органов за счет деятельности прокамбия, который откладывает кнаружи органа элементы флоэмы, внутрь - элементы ксилемы. Образовавшиеся из прокамбия проводящие пучки, состоящие из флоэмы и ксилемы, называются первичными. В результате работы прокамбия формируются два типа проводящих пучков. Если все клетки прокамбия дифференцируются в ткани проводящего пучка, то такой пучок называется закрытым. Они наблюдаются обычно у однодольных растений, он не способен к дальнейшему росту. Если между флоэмой и ксилемой остается прокамбиальный слой, дающий начало камбию, пучок продолжает расти благодаря образованию новых элементов флоэмы и ксилемы и называется открытым. Такой пучок характерен для двудольных растений, стебли и корни которых способны к утолщению.
Клетки камбия, делясь периклинально, образуют элементы вторичной ксилемы и флоэмы.
Классификация пучков. По элементарному составу различают четыре группы пучков. Простые пучки - наиболее примитивные и состоят из однородных гистологических элементов, либо из одних трахеид, либо из одних ситовидных трубок. Общие пучки - трахеиды, сосуды и ситовидные трубки — расположены бок о бок. Сложные пучки, помимо проводящих элементов, содержат паренхимные элементы. Сосудисто-волокнистые пучки, наиболее специализированные и широко распространенные, включают все элементы ксилемы и флоэмы.
Расположение ксилемы и флоэмы может быть различным, в связи с этим выделяют следующие типы проводящих пучков. Коллатеральный, или бокобочньш, когда флоэмы и ксилема располагаются бок о бок, т. е. на одном радиусе. Наружная часть пучка представлена флоэмой, внутренняя - ксилемой. Этот тип пучка наиболее распространен. Биколлатеральный, или дважды, бокобочньш, — флоэма прилегает к ксилеме с обеих сторон, при этом один участок флоэмы более мощный — наружный, другой - слаборазвитый — внутренний (семейства тыквенных, пасленовых, колокольчиковых, сложноцветных). Образуются путем слияния двух коллатеральных пучков. Концентрический встречается относительно редко. Различают амфивазальный, в котором ксилема замкнутым кольцом окружает флоэму; встречается у однодольных, например в корневище ландыша, и амфикрибраль-ный, в котором флоэма окружает ксилему, встречается у папоротниковых. В радиальном пучке участки флоэмы и ксилемы лежат по разным радиусам, разделены паренхимной тканью. Этот тип пучка характерен для первичного строения корня двудольных растений. У однодольных такие пучки сохраняются до конца жизни. У двудольных при переходе от первичного ко вторичному строению корня радиальное расположение сменяется коллатеральным.
Во всех открытых пучках, где из прокамбия дифференцируется камбий, структура динамична и постепенно усложняется. Камбий, расположенный внутри пучка, называется пучковым. Он представлен или сплошным замкнутым кольцом, или в форме тяжей, отграниченных от основной паренхимы. Работа камбия приводит к разрастанию пучков в радиальной и тангенциальной плоскости, в результате образуется межпучковый камбий. Участки межпучкового камбия, соединяясь с участками пучкового камбия, образуют камбиальное кольцо. Межпучковый камбий начинает формировать новые пучки, камбий в пучках - новые элементы ксилемы и флоэмы.
Дата добавления: 2015-01-19; просмотров: 9188;