Конвергентно-аккомодацион-но-зрачковый рефлекс
При переводе взгляда от далеко расположенного предмета к предмету, лежащему вблизи, происходит одновременно сужение зрачка, аккомодация и конвергенция. Все три компонента рефлекса экспериментально воспроизводятся стимуляцией затылочной коры головного мозга. Необходимо отметить, что конвергентно-аккомодационно-зрачковый рефлекс (рис. 4.5.9, 4.5.12, 4.5.13) не является истинным рефлексом. Изменение размера зрачка, процессы аккомодации и сведение глазных яблок являются ассоциированным движением, обеспечиваемым надъядерными связями между нейронами, обслуживающими сфинктер, ресничную мышцу и наружные мышцы глаза. Об этом свидетельствует то, что зрачок сокращается при отсутствии изменения освещенности глаза.
При рефлекторной зрачковой реакции и аккомодации глаза импульсы первоначально возникают в небольшой по размерам области сетчатки, предположительно в районе центральной ямки. Раздражителем, вызывающим аккомодационный рефлекторный ответ, служит нарушение четкой фокусировки изображения на поверхности сетчатки, которое регистрируется зрительной корой.
Из сетчатки импульс передается по зрительному нерву и зрительному тракту к наружному коленчатому телу. При этом в области зрительного перекреста волокна от носовых полей сетчатки переходят на противоположную сторону, а волокна от височных отделов сетчатки остаются неперекрещенными.
Из наружного коленчатого тела сигнал передается в первичную зрительную кору (поле 17) и далее во вторичную зрительную кору (поле 18). Из поля 18 сигналы поступают в пре-тектальную область через корково-покрышеч-
ный путь, где и образуют релейные синапсы (рис. 4.5.13).
Нейроны претектальной области устанавливают связь с ядром Якубовича—Эдингера— Вестфаля, эфференты которого являются пре-ганглионарными волокнами, идущими в составе глазодвигательного (III) нерва к ресничному ганглию [548, 549].
Ядро Якубовича—Эдингера—Вестфаля находится под ингибирующим действием ретикулярной формации моста. Усиление ингибирования приводит к расширению зрачка, а снижение — к сужению. Разрушение ингибирующих путей (например, при кровоизлияниях в области моста или сифилитическом поражении) вызывает сужение зрачка, которое сочетается с типичными клиническими признаками заболевания.
|
| 10 |
В последние годы получены данные, свидетельствующие о том, что волокна, обеспечивающие сужение зрачка при конвергентно-ак-комодационно-зрачковом рефлексе проходят несколько иным путем, чем волокна, обеспе-
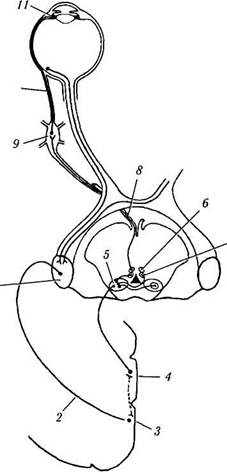
Рис. 4.5.12. Схема иннервации мышц ресничного тела:
/ — наружное коленчатое тело; 2 — зрительная лучистость; 3 — зрительная кора; 4 — перистриарная кора; 5 — претекталь-ная область; 6 — медиальная колонка висцеральных нейронов; 7 — латеральная колонка висцеральных нейронов; 8 — глазодвигательный нерв; 9 — ресничный ганглий; 10 — короткие ресничные нервы; // — мышцы ресничного тела
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ

| ......... , li-^L-VРадужка %^ С/ :Д~.— Ресничное тело J Хрусталик |
| Аккомодация |
| Сетчатка |
| Зрительный перекрест i |
| ------ 4 1 ЗК 1--------------- |
| Подушка таламуса |
 Сужение зрачка
Сужение зрачка
Ресничный ганглий
Ассоциативная кора височной, теменной и затылочной долей



 Полосатое тело
Полосатое тело
ВЛЯ
Поля 4—6
J



 КЯ
КЯ

 Мозжечок
Мозжечок
| . Мост
Рис. 4.5.13. Рефлекторные дуги зрачково-конвергентно-аккомодационного рефлекса (по Шаде, Форд, 1976):
ЯЭВ — ядро Якубовича—Эдингера—Вестфаля; НКТ — наружное коленчатое тело; ПрТ — претектальная область; ВБ — верхние бугорки четверохолмия; ЗК — задняя комиссура; ДЛРФ — дорзолатеральная ретикулярная формация; РФСМ — ретикулярная формация среднего мозга; ВЛЯ — вентролатеральное ядро зрительного бугра; КЯ — красное ядро
Вегетативная (автономная) иннервация глаза
 чивающие зрачковый («световой») рефлекс. Полагают, что волокна конвергентно-аккомода-ционно-зрачкового рефлекса приближаются к претектальному ядру с вентральной стороны. Этим можно объяснить потерю реакции зрачка при освещении глаза прежде, чем разобщается конвергентно-аккомодационно-зрачковый рефлекс. Подобное состояние встречается при дор-зальных инфильтративных (сдавливающих) повреждениях покрышки среднего мозга на уровне верхнего двухолмия (синдром Парино).
чивающие зрачковый («световой») рефлекс. Полагают, что волокна конвергентно-аккомода-ционно-зрачкового рефлекса приближаются к претектальному ядру с вентральной стороны. Этим можно объяснить потерю реакции зрачка при освещении глаза прежде, чем разобщается конвергентно-аккомодационно-зрачковый рефлекс. Подобное состояние встречается при дор-зальных инфильтративных (сдавливающих) повреждениях покрышки среднего мозга на уровне верхнего двухолмия (синдром Парино).
Аксоны нейронов ядра Якубовича—Эдинге-ра—Вестфаля поступают затем в ресничный ганглий, оканчивась синапсами на клетках обоих его компонентов. Некоторые волокна заканчиваются на нейронах, иннервирующих ресничное тело, и вызывают сокращение его мышечных волокон. В результате этого уменьшается напряжение связки, поддерживающей хрусталик, что и приводит к изменению фокусного расстояния. Именно благодаря этому на сетчатке формируется четко сфокусированное изображение.
Другие преганглионарные волокна образуют связи с частью ресничного узла, обеспечивающей сужение зрачка.
Третьим компонентом реакции аккомодации является конвергенция глаз при рассматривании близко расположенных предметов. Первоначально полагали, что центры конвергенции находятся в среднем мозге, однако существование таких центров не было доказано. В настоящее время считается, что эта реакция осуществляется при участии корковых связей [548]. Так, предполагают, что при этом реализуется связь между полем 18 зрительной коры с подушкой зрительного бугра (pulvinar thalami), которая, в свою очередь, посылает волокна в ассоциативные центры височной, теменной и затылочной долей.
Эти центры могут быть связаны с ретикулярной формацией среднего мозга, верхними бугорками и дорзолатеральными отделами ретикулярной формации. Благодаря таким связям с ретикулярной формацией сигналы могут идти к нейронам ядра глазодвигательного нерва, ин-нервирующим внутреннюю прямую мышцу глаза, а также к ядру отводящего нерва с целью торможения клеток, иннервирующих наружную прямую мышцу глаза.
Вторая возможная система связей может идти через полосатое тело к вентролатераль-ным ядрам зрительного бугра (nuclei ventro-laterales {thalami)). От этих ядер связи направляются к 4-му и 6-му полям коры головного мозга (моторная и премоторная области). Из этих полей коры импульсы через дорзолате-ральные отделы ретикулярной формации могут достигать ядер черепно-мозговых нервов. В этом варианте возможны также связи через ядра моста с мозжечком и связи с красным ядром, что делает реальной координирующую роль мозжечка в реакции конвергенции.
Как указано выше, при конвергентно-акко-модационно-зрачковом рефлексе происходит фиксация, которая, по существу, складывается из аккомодации и сведения глазных яблок. Некоторые исследователи предполагают, что рефлекс начинается с конвергенции. При этом про-приоцептивные импульсы от внутренней прямой мышцы направляются через глазодвигательный нерв или первую ветвь тройничного нерва к ядру среднемозгового пути тройничного нерва (nucleus mesencephalicus nervi trigemi-nalis (п. mesencephalicus trigemini)). Отсюда импульс направляется к ядру глазодвигательного нерва. После оставления глазодвигательного нерва импульс проходит через ресничный ганглий и проходит к сфинктеру радужки.
4.5.7. Мигательный рефлекс
Этот рефлекс был описан в 1896 г. Overend. Сводится рефлекс к сокращению круговой мышцы глаза при механическом или электичес-ком раздражении верхнего глазничного нерва первоначально со стороны раздражения, а несколько позже с двух сторон [157, 305]. Рефлекторная дуга замыкается на уровне ядер ствола мозга благодаря наличию интернейронов. Именно по этой причине возникает патологическое сокращение круговой мышцы глаза при заболеваниях ствола мозга, таких как синдром Валленберга [158].
Дата добавления: 2015-03-26; просмотров: 1633;