Анатомия головного мозга. Слой пирамидных клеток варьирует по ширине и состоит из клеток конической формы
 Слой пирамидных клеток варьирует по ширине и состоит из клеток конической формы. Максимально выражен он в ассоциативных и сенсомоторных областях коры. На апикальной поверхности нейронов располагаются дендриты и аксоны. В этом слое обнаруживаются также многочисленные вставочные нейроны (интернейроны) звездчатой формы, нейроны с отростками, ориентированными как в вертикальной плоскости (веретенообразные клетки), так и горизонтальной (корзинкоподобные клетки) плоскости. Их аксоны и дендриты распространяются и на другие слои коры. Слой выполняет преимущественно ассоциативные функции.
Слой пирамидных клеток варьирует по ширине и состоит из клеток конической формы. Максимально выражен он в ассоциативных и сенсомоторных областях коры. На апикальной поверхности нейронов располагаются дендриты и аксоны. В этом слое обнаруживаются также многочисленные вставочные нейроны (интернейроны) звездчатой формы, нейроны с отростками, ориентированными как в вертикальной плоскости (веретенообразные клетки), так и горизонтальной (корзинкоподобные клетки) плоскости. Их аксоны и дендриты распространяются и на другие слои коры. Слой выполняет преимущественно ассоциативные функции.
IV. Внутренний зернистый слой (внутрен
няя зернистая пластинка; lamina granularis
inter na).
Этот слой имеет различную толщину. Наиболее широкий он в области зрительной и слуховой коры мозга. Состоит слой, главным образом, из вставочных нейронов звездчатой формы и небольшого количества пирамидных клеток. В вертикальной плоскости в нем распространяются многочисленные аксоны и дендриты. Выявляются «уплотнения», состоящие из горизонтально распространяющихся отростков, так называемая наружная связка Бейларгера (Baillarger), которая наиболее выделяется в первичной зрительной коре. Слой подразделен на подслои IVA, IVB и IVC (см. «Зрительный путь»). В этом слое заканчивается основная масса таламических афферентных волокон. Аксоны клеток слоя образуют связи с клетками выше- и нижележащих слоев коры.
V. Слой ганглиозных клеток (внутренняя
пирамидная пластинка; lamina pyramidalis in-
terna (ganglionaris)).
Слой ганглиозных клеток также содержит звездчатые и крупные пирамидные клетки (гигантские клетки Беца). Определяются в составе 4-го и б-го полей (по Бродману). Подобно другим слоям коры в этом слое между клетками распространяются дендриты и аксоны нейронов. Аксоны гигантских и крупных пирамидных клеток проецируются на ядра головного и спинного мозга. Наиболее длинные из них достигают каудальных сегментов спинного мозга. В V слое сосредоточено большинство корковых проекционных эфферентов.
VI. Полиморфный, или веретенообразный,
слой (мультиформная пластинка; lamina multi-
formis).
Последний слой граничит с белым веществом головного мозга. Состоит он из маленьких нейронов, главным образом «зернистых», вере-теновидных, звездоподобных вставочных нейронов. Встречаются и маленькие пирамидные клетки. Некоторые из пирамидных клеток (нейроны Мартинотти) отдают длинный двигательный аксон в молекулярный (плексиформный) слой, а вертикально ориентированные его дендриты разветвляются в более глубоких слоях
коры. Большинство отростков вставочных нейронов и пирамидных клеток отдают аксоны, которые оставляют кору и направляются к ба-зальным ганглиям, зрительному бугру, гиппо-кампу, ядрам ствола головного и спинного мозга. В этом слое обнаруживаются различные типы синапсов — аксо-дендритные, аксо-сома-тические, аксо-аксонные [540].
Необходимо еще раз подчеркнуть, что типы клеток, их количество и свойственные им связи отличаются в различных участках коры. Это относится и к первичной зрительной коре, о чем более подробно будет изложено в соответствующем разделе (см. «Зрительный путь»).
Классификации неокортекса.Обычное шес-тислойное строение коры несколько изменяется в различных участках мозга. Эти структурные различия строения по площади коры довольно подробно изучены, и на этой основе созданы цитоархитектонические схемы строения коры, отражающие ее функциональные особенности. Наиболее распространенной классификацией является классификация Бродмана (рис. 4.1.9). В соответствии с ней всю поверхность коры можно разделить на 52 поля (области). Эта классификация получила широкое распростра-
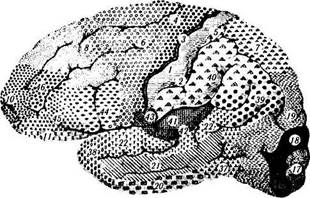
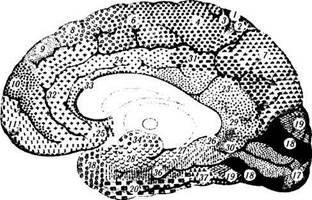
Рис. 4.1.9.Цитоархитектоническая карта коры головного мозга по Бродману:
а — наружная поверхность; б—внутренняя поверхность
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
нение в связи с тем, что структурные различия хорошо коррелируют с функциональными особенностями, о чем будет изложено несколько ниже.
|
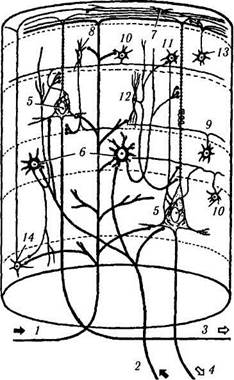
Вертикальная организация коры. Общий поток восходящей и нисходящей информации к коре и от нее организован вертикально. То есть информация, идущая от зрительного бугра, передается звездчатым клеткам IV слоя, которые, в свою очередь, передают ее более высоко и более низко расположенным клеточным слоям для дальнейшей обработки. Подобная вертикальная, или колоночная, организация довольно легко демонстрируется при использовании не только новых морфологических, но и физиологических методов (рис. 4.1.10). Эти методы позволили точно установить, что кору мозга можно подразделить на структурно-функциональные колонки, распространяющиеся от мягкой мозговой оболочки до белого вещества [3, 28, 286]. При этом в границах колонки все нейроны выполняют одну и ту же четко определенную функцию. Ширина такой функциональной ко-
III IV
V VI
Рис. 4.1.10. Колоночная организация коры полушарий большого мозга (по В. J1. Быкову, 1997). На схеме показаны главные ассоциативные связи клеток основных типов:
/ — кортико-кортикальное афферентное волокно; 2 — таламо-кор-тикальное афферентное волокно; 3 — ассоциативное эфферентное волокно; 4 — проекционное эфферентное волокно; 5 — пирамидная клетка; б — шипиковая звездчатая клетка; 7 — горизонтальная клетка Кахала; 8 — аксо-аксонная клетка; 9 — клет-ка-«канделябр», 10—корзинчатая клетка; //—колонковая кор-зинчатая клетка; 12 — клетка с двойным букетом дендритов; 13 — клетка с аксональным пучком; 14 — клетка Мартинотти. Римскими цифрами обозначены слои
лонки колеблется от 250 до 500 мкм, а их количество равняется 2—3 млн. Каждая колонка содержит примерно 5000 нейронов. Внутри колонки выделяют также более мелкие мини-колонки, включающие структуры, непосредственно окружающие апикальные дендриты пирамидных клеток. Колонка включает:
1) афферентные пути;
2) систему локальных связей;
3) эфферентные пути.
Афферентные пути организованы следующим образом. В центре колонки проходит примерно 100 возбуждающих кортико-кортикаль-ных волокон — аксонов пирамидных клеток других колонок данного и противоположного полушарий. Они образуют окончания во всех слоях колонки (в том числе на клетках Мартинотти, шипиковых звездчатых клетках, латеральных дендритах пирамидных клеток) и проходят до I слоя, где образуют ветви, уходящие за ее пределы.
Специфические афферентные импульсы по таламо-кортикальным волокнам поступают на тела и дендриты пирамидных клеток и на шипи-ковые звездчатые клетки IVDcnoH (последние по своим аксонам передают их на апикальные и базальные дендриты пирамидных клеток).
Система локальных связей формируется вставочными нейронами колонки, которые включают более десятка типов клеток. Часть из них обладает тормозной функцией и регулирует преимущественно активность пирамидных клеток. Из тормозных нейронов колонки наибольшее значение имеют следующие [3] (см. рис. 4.1.10):
1. Аксо-аксонные клетки, тела которых ле
жат во II—III слоях, а аксоны идут горизон
тально, отдавая многочисленные терминальные
веточки, которые образуют тормозные синапсы
на начальных сегментах аксонов пирамидных
клеток II и III слоев.
2. Клетки-«канделябры», которые встреча
ются во всех внутренних слоях коры. Их аксон-
ные коллатерали идут горизонтально и дают
несколько восходящих и нисходящих веточек,
которые образуют спиральные ветвления во
круг апикальных дендритов пирамидных клеток.
3. Корзинчатые клетки, которые располага
ются во II слое, на границе III и IV, а также
IV и V слоев. Их аксоны проходят горизонталь
но на расстояние до 3 мм и, оплетая тела круп
ных и средних пирамидных клеток, влияют на
20—30 соседних колонок. Колонковые корзин
чатые клетки обеспечивают торможение пира
мидных клеток по вертикали внутри данной
колонки.
4. Клетки с двойным букетом дендритов, от
ходящих вертикально от полюсов клетки, рас
положены во II—III слоях. Их аксон дает кол
латерали, образующие контакты с дендритами
как пирамидных клеток, так и непирамидных (в
том числе тормозных) нейронов. Первый тип
контактов опосредует угнетение пирамидных
Анатомия головного мозга
 клеток, а второй — их активацию путем снятия торможения.
клеток, а второй — их активацию путем снятия торможения.
5. Клетки с аксонным пучком (кисточкой) — звездчатые нейроны II слоя, аксоны которых ветвятся в I слое, образуя связи с дистальными сегментами апикальных дендритов пирамидных клеток и с горизонтальными ветвями кортико-кортикальных волокон.
Эфферентные пути формируются следующим образом. Аксоны средних пирамидных клеток III слоя устанавливают связи преимущественно с соседними колонками и колонками противоположного полушария, а аксоны крупных и гигантских пирамидных клеток V слоя, помимо этого, направляются в подкорковые центры, образуя вместе с аксонами веретено-видных клеток VI слоя систему эфферентных волокон коры.
Определенные особенности вертикальной организации коры существуют в области зрительной коры. Эти сведения будут приведены в разделе, посвященном зрительной коре.
Приводящие и отводящие связи коры.Нейроны коры можно подразделить на ряд категорий в соответствии с особенностями их связей. К первой категории нейронов относятся проекционные нейроны, которые передают импульсы к подкорковым центрам, таким как зрительный бугор, ствол мозга, спинной мозг или базальные ганглии. Ко второй категории относятся вставочные нейроны, формирующие связи с другими нейронами в том же самом полушарии. К третьей категории относятся ко-миссурные нейроны, которые напоминают по
функции вставочные нейроны за исключением того, что они посылают аксоны в противоположное полушарие через мозолистое тело или переднюю спайку мозга [3, 4, 6, 8].
Большинство афферентов заканчивается в
IV слое коры головного мозга (рис. 4.1.11).
Эфференты, исходящие из коры, начинаются от
нейронов V и VI слоев. Пирамидные нейроны
V слоя проецируются непосредственно на ствол
мозга или спинной мозг. Нейроны VI слоя явля
ются основным источником прямых проекций
на зрительный бугор. Эфференты, направляю
щиеся к другим областям коры (т. е. вставоч
ные нейроны), исходят от нейронов IIIи VI
слоев.
Архикортекс.Филогенетически наиболее старые структуры коры названы палеокортек-сом, мезокортексом и архикортексом. Они характеризуются слоистой организацией, но число слоев нейронов в этих участках коры меньше шести. Основной областью палеокортекса является обонятельная кора, мезокортекса — кора поясной извилины, а основной частью архикортекса — гиппокамп. Гиппокамп отличается от коры обратным расположением слоев. При этом серое вещество гиппокампа располагается внутри, а белое — снаружи. Часть гиппокампа состоит только из трех слоев клеток.
Нейромедиаторная система коры.В коре выявлено более десятка различных нейроме-диаторов и рецепторов к ним. Это норадрена-лин, серотонин, ацетилхолин, вазоинтерстици-альный кишечный полипептид (VIP), соматоста-тин, холецистокинин, глютамат и др.
Слои
Афференты
Эфференты
Мягкая оболочка
 III
III
А
А
Кора мозга
IV

 VI
VI
А
 Белое вещество
Белое вещество


 V V
V V
Кортикокор-тикальные
Кортикокор-тикальные
Таламокор-тикальные
Кортикотала- Кортикобульбарные мические Кортикопонтинные
Кортикоретикулярные
Кортикоспинальные
Кортикостриарные
Рис. 4.1.11. Особенности связей коры головного мозга (объяснение в тексте)
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ

|
| IV |
Функции коры
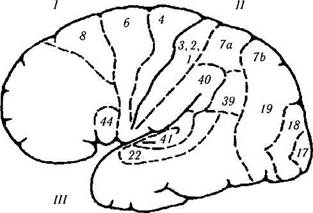
Лобная доля. Основной функцией лобных долей является контроль произвольных движений, координация двигательных механизмов выразительной речи, а также обеспечение «творческого», или абстрактного, мышления.
Лобную долю можно разделить на пять областей: моторную, премоторную, лобную моторную зрительную, моторную область речи Бро-ка и лобную ассоциативную области (рис. 4.1.12, 4.1.13).
Первичная двигательная область (моторная область; 4-е поле по Бродману).
Моторная область находится впереди центральной борозды, отделяющей лобную долю от теменной доли. Состоит она из большого количества гигантских пирамидных клеток Беца.
|
| IV |
| III |
В ней представлены движения, а не индивидуальные мышцы. Клетки, управляющие движениями, расположены таким образом, что движения нижней половины тела представлены в верхней части извилины, а верхней — в нижней. Аксоны нейронов спускаются через внутреннюю капсулу и ножки мозга в виде кортико-бульбарных и кортикоспинальных путей.
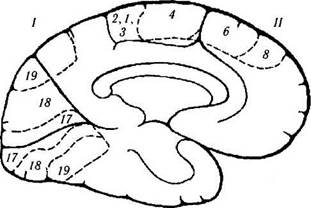
Рис. 4.1.12. Схематическое изображение функций коры большого мозга, локализованных на медиальной поверхности, и их нарушение при различных заболеваниях, сопровождающихся возбуждением или разрушением коры (по Crosby et al., 1962 [ ]):
/ — теменная доля. Центральная извилина (4). Левая затылочная область: —алексия без аграфии. Задняя кора: —сложность перевода взгляда; —синдром Балинта (двухсторонний). // — лобная доля. Дополнительное двигательное поле (6): +поднятие противоположной руки, поворот глаз и головы к руке; + сокращение зрачка; + задержка речи. Лобное двигательное глазное поле (8). Поясная извилина: + расширение зрачка; + слезоотделение. Орбитальная поверхность. Ill — затылочнаядоля. Первичная зрительная кора. Контрлатеральное зрение: —правосторонняя гемианопсия. IV—височная доля:паралич движений глаз; ипсилатеральная гемиплегия; гидроцефалия; децеребральная регидность; педункулярный галлюциноз; контрлатеральная гемианопсия; синдром Паринауда. Под
поверхностью: — распознание лица.
Примечание. Цитоархитектонические поля соответствуют классификации Бродмана. Плюс обозначает результат электростимуляций коры, а минус — результат разрушения поля
Рис. 4.1.13. Латеральная поверхность мозга (обозначения аналогичны приведенным на предыдущем рис.):
/ — лобная доля. Двигательное поле (4). Премоторное поле (6). Двигательное глазное поле (8): + контрлатеральная сак-када; + закрытие противоположного глаза; + гомолатеральное содружественное отклонение глаза. Двигательное речевое поле (44): —двигательная (моторная) афазия. // — теменная доля:-(-простые галлюцинозы; —иллюзии; — метаморфопсия; —потеря внимания; —агнозия; — апраксия; —ослабление оптокинетического нистагма; — нижняя квадрантная гемианописия. Супрамар-гинальная извилина (40): —амнезия; —жаргонная афазия. Угловая извилина (39): —аграфия; — акалькулия; —утеря способности различать левую и правую сторону; пальцевая агнозия. Соматическая чувствительность. ///—височная доля:+ зрительные галлюцинации; + deja vu; + нарушение обоняния; + припадки; + автоматизмы; —верхняя квадрианопсия; —потеря памяти; — синдром Клювер—Бюси. Первичная слуховая кора. Вторичная слуховая кора: —слуховая афазия. Planum temporale: — дислексия. IV—затылочная доля (17, 18, 19): + ощущение света; -(-ощущение цветов; -(-ощущение движения; + ощущение линейных изображений. Зрительные ассоциативные поля (18, 19): -(-контрлатеральные движения глаза типа плавного преследования; + вергентные движения; —синдром Антона
Удаление двигательной области коры головного мозга ведет к параличу.
Премоторная область (поле 6) лежит непосредственно кпереди первичной двигательной коры, занимая заднюю часть трех горизонтально ориентированных лобных извилин.
Формирует она волокна фронтопонтомоз-жечковых трактов, а также отдает многочисленные волокна базальным ганглиям.
Предполагают, что премоторная кора вовлечена в планирование двигательных действий. Об этом свидетельствует то, что лишь при размышлении о предполагаемом действии нейроны шестого поля возбуждаются. Стимуляция этих нейронов приводит к подъему руки противоположной стороны, а голова и глаза поворачиваются в направлении поднятой конечности. Зрачки расширяются, и нарушается речь (см. рис. 4.1.12, 4.1.13).
Разрушение этой области сопровождается псевдомозжечковой атаксией противоположной стороны. Нарушается также возможность копирования изученных движений (апраксия). При этом отдельные движения не повреждены, но больной не способен производить их в нужной последовательности.
Анатомия головного мозга
 Лобная моторная зрительная область (поле 8 по Бродману).
Лобная моторная зрительная область (поле 8 по Бродману).
Эта область (поле 8 и частично 6 и 9) занимает заднюю часть медиальной лобной извилины (gyrus frontalis medius).
Лобная моторная зрительная область важна в произвольных движениях глаза. Некоторые участки определяют конвергенцию, дивергенцию и аккомодацию [96, 322].
Необходимо отметить, что движение глаз, зрачка и век могут выявляться при электростимуляции почти всех областей полушарий мозга [276]. Электростимуляция лобной моторной глазной области приводит к саккадическим движениям, в отличие от реакции при стимуляции полей 18, 19 и 22 (плавное слежение и вергентные движения). Стимуляция 8-го поля у обезьян сопровождается содружественным контрлатеральным отклонением глаз. При этом расширяется зрачок, а голова и туловище поворачиваются в противоположную сторону. Повреждение лобной доли вызывает обратимую потерю способности к контрлатеральным саккадическим движениям [135].
Эта область коры отличается многочисленными связями.
Афферентные волокна поступают в зрительную лобную кору от зрительного бугра, а также от верхне-височной, перистриарной и теменной (область 7) коры. Здесь обнаружены нейроны, контролирующие движения глаз.
Эфференты проецируются на базальные ганглии, зрительный бугор, претектальную область (включая ядро зрительного тракта), верхние бугорки четверохолмия и часть ретикулярной формации моста [240, 321, 447].
В свою очередь необходимо отметить, что лобные зрительные области проецируются на контрлатеральные лобные поля и на области коры с этой же стороны. Обеспечивают они зрительное восприятие (на 7-м поле теменной коры и перистриатной коре).
При проекции на средний мозг волокна пре-лобной коры пересекают переднюю часть внутренней капсулы и делятся на дорзальные и вентральные пути в ростральной части промежуточного мозга [330].
Дорзальный трансталамический путь пересекает дорзомедиальное и интраламилярное ядро зрительного бугра и среднюю часть подушки, образуя при этом небольшое количество синаптических связей. Синапсы обнаруживаются также в претектальных ядрах и в верхних бугорках четверохолмия.
Вентральный педункулотегментальный путь проходит в наиболее вентральной части ножки мозга, поставляя волокна к ядрам гипоталамуса, а глубже — к верхним бугоркам четверохолмия. При этом лобные зрительные области обладают топографической проекцией на верхние бугорки четверохолмия [446]. Завершается путь в ретикулярной формации моста [240].
Третья префронтальная связка возникает в области диэнцефально-мезенцефалического соединения, расположенного около красного ядра. Этот путь проецируется на ядро медиального продольного пучка и промежуточное ядро Кахаля с этой же стороны, а также на ядро Даркшевича и растральную часть глазодвигательного ядра, с обеих сторон.
Различные участки лобной зрительной коры контролируют саккадические движения глаза различной амплитуды, а также направление сак-кад. Они обеспечивают так называемое «зрительное внимание», направляя глаза к «цели» [119, 172].
В настоящее время выясняется роль лобной доли в функционировании век. Произвольное закрытие век требует расслабления леватора век и сокращения круговой мышцы глаза. Существуют разногласия относительно локализации участков мозга, контролирующих волевые движения века. Эти участки могут располагаться в двигательной премоторной или лобной моторной зрительной областях. Стимуляция этой области ведет к контрлатеральному закрытию века. Односторонние повреждения недоминирующих лобных долей могут сопровождаться двусторонним нарушением закрытия века.
Моторная речевая область Брока (поля 44 и 45 по Бродману).
Эта область располагается в задне-нижней части лобной доли [3, 4, 6, 8]. Она вносит вклад в инициирование речи. Повреждение области Брока приводит к афазии, заключающейся в сложности соединения в определенной последовательности слов для обеспечения связанной речи (экспрессивная афазия). Больные при этом понимают как письменную, так и разговорную речь, но обычно не могут ничего произнести. Нарушена также способность писать, хотя рука может использоваться для выполнения других действий.
Лобная ассоциативная область. Эта важная область в передней половине лобной доли ответственна за инициирование всех типов по-веденческо-интеллектуальных, висцеральных и эмоциональных функций. Изменения памяти, интеллекта и индивидуальности наступают при раздражении или массивном разрушении лобных долей. Это особенно выражено в тех случаях, когда поражено доминирующее полушарие.
Обонятельный тракт находится ниже глазничной поверхности лобной доли и является единственным сенсорным путем, достигающим коры без образования синапсов в зрительном бугре. Односторонняя опухоль глазничной поверхности лобной доли (менингиома обонятельной борозды или менингиома в области малого крыла клиновидной кости) может привести к сдавлению обонятельного тракта и зрительного нерва. В результате развивается аносмия (отсутствие обоняния), ипсилатеральная атрофия
Глава 4. ГОЛОВНОМ МОЗГ И ГЛАЗ
 зрительного нерва и отек диска зрительного нерва с противоположной стороны (синдром Кеннеди Фостера).
зрительного нерва и отек диска зрительного нерва с противоположной стороны (синдром Кеннеди Фостера).
Поражение структур лимбической системы в пределах лобной доли, особенно поясной извилины, может вызвать нарушения эмоциональной стороны жизнедеятельности, а также висцеральные признаки, включая беспричинное слезотечение и изменение реакции зрачка на световые стимулы.
Теменнаядоля (см. рис. 4.1.12, 4.1.13). В целом функция теменных долей связана с чувствительностью, памятью, относящейся к речи и обучению, а также с ориентацией организма в пространстве путем обработки зрительной информации [17, 116, 126, 234, 558].
В переднем отделе теменной доли находится извилина, расположенная параллельно прецен-тральной извилине или двигательной области коры. Это постцентральная извилина, или первичная соматосенсорная кора (поля 3, 1 и 2) (см. рис. 4.1.13). Оставшуюся часть теменной доли можно разделить на две дольки — верхнюю и нижнюю. Поля 5 и 7, расположенные на поверхности верхней теменной дольки, связаны с корреляцией соматосенсорной информации, дающей возможность сознательной оценки веса, свойств поверхности, размера и формы предмета.
Нижняя теменная долька (супрамаргиналь-ная и угловая извилины) тесно связана с речевыми механизмами, поскольку разрушение этой области коры в левом полушарии ведет к нарушению восприятия речи.
Роль теменной доли в пространственной ориентации пока еще не изучена. Однако у больных с ее поражением отмечается нарушение этой функции.
Через теменную долю проходят приводящие волокна, соединяющие области 18 и 19 с лобными долями и стволом мозга. Эти пути включают верхний и нижний продольный пучки.
Предполагают, что теменная кора (поле 7) обеспечивает функцию «внимания» к зрительным целям [29, 86, 127, 387, 488, 501, 502].
Седьмое поле получает афференты от поясной извилины (g. cinguli), от претектальной области и верхних бугорков четверохолмия через ядра зрительного бугра. Обнаружены также афференты, идущие от лобной зрительной области и престриарной коры к коре теменной дольки.
Эфференты проецируются на претектальную область, верхние бугорки четверохолмия, серое вещество, расположенное вокруг сильвиевого водопровода, и на теменные доли двух полушарий мозга [447].
При движении глаз нейроны нижней теменной доли активизируются, что свидетельствует об их роли в контроле движений глаз, подчиняющихся воле. Активность продолжается на всем протяжении наблюдения за целью.
Клетки 7-го поля не определяют саккадичес-кие движения, поскольку их активность при саккадах не увеличивается [86, 502].
Стимуляция угловой извилины (g. angula-ris) приводит у экспериментальных животных к содружественному отклонению глаз. Это поля 38 и 39, переходящие вокруг заднего края средней височной извилины (g. temporalis medius). Оперативное удаление этой области коры не приводит к нарушению движения глаз. Двухсторонние повреждения теменной коры могут привести к развитию моторной апраксии глаза [113].
Поражение теменной доли сопровождается также нарушением зрительных моторных механизмов, обеспечивающих оптокинетический нистагм, в течение которого глаза производят саккады для поиска и фиксации цели.
Другие моторные признаки изолированных повреждений теменной доли включают содружественное отклонение глаз в противоположную сторону от места повреждения при принудительном закрытии век, малую подвижность больного и плохой контроль (непонимание) конечностей противоположной стороны. Трудности в осмыслении характера действия конечностей ведут к конституциональным и глазодвигательным апраксиям [153]. У больных с повреждениями теменной доли могут возникать трудности в процессе перемещения взгляда от одного зрительного объекта к другому, в пределах контрлатеральной половины поля зрения.
Другим признаком дисфункции теменной доли является неспособность фиксировать зрительный объект.
Кроме сенсорных функций, которые локализуются в задней губе центральной борозды Роландо, теменная доля является участком сложного процесса узнавания и осмысления. Диффузное повреждение теменных долей завершается агнозиями, зрительным невниманием, метаморфопсией, фотопсией, галлюцинациями, иллюзиями и алексией (см. рис. 4.1.12, 4.1.13).
Более подробные сведения о последствиях нарушения функции теменной доли приведены в неврологической литературе. Здесь мы приводим лишь некоторые синдромы.
Зрительная агнозия, которая часто развивается при нарушении кровообращения в ЦНС, характеризуется нарушением процесса узнавания. Агнозия может быть полной или частичной. Зрительная агнозия диагностируется в тех случаях, когда больной может идентифицировать объект только тактильно. Часто нарушено узнавание и осмысление зрительных изображений и цветового смысла изображения.
Двухсторонние повреждения теменной доли (чаще встречается при старческом слабоумии, размягчении височно-затылочной и теменной областей коры, ранениях задних отделов обоих полушарий, энцефалите) сопровождаются зри-
Анатомия головного мозга
 тельной агнозией и зрительной атаксией (синдром Балинта).
тельной агнозией и зрительной атаксией (синдром Балинта).
Зрительная пространственная агнозия характеризуется возникновением трудностей при оценке размера объекта и расстояния до него. Отмечается также появление неправильного решения «синтетических» пространственных задач типа чтения карты. Симультантная агнозия сводится к тому, что у больных сохранена способность узнавания отдельных объектов, но отсутствует способность воспринимать группу объектов (изображений) как целое.
Астерогнозия характеризуется тем, что больной может анализировать объекты по их виду, но не способен охарактеризовать их тактильно. Это развивается вследствие нарушения сложного взаимодействия между тактильной и зрительной информацией в теменных долях коры.
Диффузные поражения теменных долей могут привести к возникновению иллюзий и мета-морфопсий. Эти искажения включают микро-псию, макропсию. Больные отмечают движение стационарных целей, неправильно оценивают расстояние до объектов. К специальным типам иллюзий относятся палинопсия (нарушение зрительного восприятия в виде сохранения или повторного возникновения зрительного образа после исчезновения предмета из поля зрения), полиопия галлюцинаторная и др.
Иллюзии развиваются в результате диффузных нарушений коры, а галлюцинации и фото-псии — при развитии опухоли или токсическом повреждении коры.
Особенности патологии теменной доли могут зависеть и от того, локализуется ли патологический очаг в доминирующем (обычно левом) или недоминирующем полушариях. Доминирующая теменная доля ответственна за развитие нарушения речи типа слуховой сенсорной афазии, зрительной агнозии и аграфии.
При повреждении доминирующей угловой извилины (чаще в результате нарушения кровообращения в бассейне угловой артерии) развиваются нистагм, зрительно-агностические расстройства, аграфия, акалькулия (неспособность складывать или вычитать цифры), пальцевая агнозия, невозможность различать правую и левую стороны (синдром Герстманна; синдром угловой извилины). Возможно развитие гомо-нимной гемианопсии.
При повреждении недоминирующей угловой извилины нарушается взаимодействие больного с окружающей средой. В результате развивается зрительная дезориентация и топографическая агнозия.
Кпереди от угловой извилины и поперек надвисочной борозды находится надкраевая извилина. В доминирующем полушарии эта область является местом встречи зрительных и слуховых компонентов языка [276, 442, 621].
Повреждение задних теменных ассоциативных областей нередко завершается развитием
апраксии (трудность выполнения «тонких» специализированных движений). Это, по-видимому, связано с недопониманием необходимой последовательности движений для выполнения той или иной функции (т. е. больному неясно, что и в какой последовательности должно быть выполнено). У больного отсутствует способность нарисовать простую диаграмму (конструктивная апраксия). Менее серьезной формой этой патологии является осязательная агнозия (неспособность распознать объект при пальпации). Иногда это заболевание называется асте-рогнозией. При этом нет никаких признаков потери осязательной или проприоцептивной чувствительности, а нарушена интеграция зрительной и соматосенсорной информации. Апраксия и астерогнозия чаще развиваются после повреждения левого полушария головного мозга. Астерогнозия обычно ограничена противоположной стороной тела, в то время как апраксия обычно двухсторонняя.
Зрительная и слуховая информация жестко связана с функцией языка. По этой причине повреждение полей 39 и 40, также как и части слуховой ассоциативной области, обычно приводит к афазии, известной как афазия Вер-нике (Wernicke). Если повреждение ограничено только угловой извилиной, теряется способность понимать письменный язык (алексия) и способность писать (аграфия). При этом больные понимают речь. Это состояние обычно развивается после травмы.
Интеграция зрительной и соматосенсорной информации важна для формирования «образа тела», т. е. понимания особенностей положения тела в пространстве. Обширные повреждения полей 7, 39 и 40 одного полушария могут привести к развитию «непонимания» или «пренебрежения» противоположной половины тела. Больные не в состоянии перевязать или разбинтовать собственные конечности. Они также не узнают собственные конечности.
Затылочная доля (поля 17, 18 и 19). Вдоль шпорной борозды (fissura calcarina) располагается первичная зрительная область (поле 17). К ней примыкает «оценочная» зрительная область (поле 18), которая, как полагают, преобразует сигналы, получаемые полем 17, в зрительные образы.
Поле 17 затылочной коры является зоной приема и обработки зрительной информации и обладает важными связями с двигательными зрительными участками коры. Именно на этом уровне зрительной коры оценивается значение увиденного [395]. Раздражение этой области вызывает галлюцинации и образы типа сновидений.
Двигательные связи затылочной доли заслуживают внимания и потому, что они вовлечены в оптокинетический нистагм, рефлекс аккомодации и регуляции бинокулярного зрения.
Поля 18 и 19 взаимозависимы и имеют большее количество связей с остальной частью моз-
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 га. Предполагают, что область 18 больше заинтересована объединением зрительной информации, полученной в 17-м поле, в то время как поле 19 переводит эту информацию в более сложную мысленную и двигательную деятельность (см. «Зрительный путь»).
га. Предполагают, что область 18 больше заинтересована объединением зрительной информации, полученной в 17-м поле, в то время как поле 19 переводит эту информацию в более сложную мысленную и двигательную деятельность (см. «Зрительный путь»).
Последние анатомические и физиологические исследования выявили, что существуют, по крайней мере, 10 подобластей полей 18 и 19, нейроны которых обладают способностью реагировать на зрительные стимулы. Каждая из этих подобластей связана с 17-м полем. При этом между собой они не связаны. Повреждение этих подобластей не приводит к слепоте. Нарушаются некоторые зрительные функции, например анализ цвета, восприятие цельности объекта и др.
Зрительная ассоциативная кора, расположенная в некоторых областях височной доли, также вовлечена в обработку зрительной информации. В наибольшей степени это относится к полю 21 (в средней височной коре). Именно это поле обеспечивает узнавание образа, а ее повреждение завершается агнозией.
Выше приведенные свойства зрительной коры обеспечиваются и многочисленными горизонтальными и вертикальными связями между нейронами различных слоев зрительной коры. Именно благодаря их наличию отмечается исключительная пластичность зрительной коры при обработке зрительной информации различного типа [69, 94, 162, 197, 199, 352, 486].
Более подробно структурная организация зрительной коры и ее функции будут приведены в следующем разделе.
Височная доля(поля 41, 22) (см.рис. 4.1.8, 4.1.12, 4.1.13).
Эта часть мозга связана с восприятием слуховых ощущений, участвует в речевой функции посредством слухового контроля речи, а также играет определенную роль в оценке пространства и в функции памяти. Давно установлено, что поперечная височная извилина (поле 41) является первичным слуховым центром. Небольшое поле (22), окружающее эту извилину, рассматривают как «психический», или «оценочный», слуховой центр. Раздражение большей части височной доли, особенно вдоль средней височной извилины, вызывает ощущения, которые больной описывает как слуховые. Больные с поражением «психического поля» височной доли иногда утрачивают способность оценки звуковых тонов. При этом больной не узнает мотивов, а музыкальные звуки воспринимаются им как беспорядочный шум.
Поле 22 левого полушария также связывают с речевой функцией, поскольку поражение этой области приводит к потере способности понимать значение слов. Существуют также данные, свидетельствующие о том, что височная доля связана с «вестибулярной» чувствительностью (чувство равновесия), так как
стимуляция задних отделов верхней височной извилины у больного, находящегося в сознании, вызывает головокружение и ощущение вращения.
Глубокие поражения височной доли часто затрагивают самые нижние волокна зрительной лучистости, идущие от наружного коленчатого тела. При этом нарушены функции зрения. Чаще встречается верхняя квадрантная гемиа-нопсия.
При поражении височной или психомоторной доли развивается эпилепсия, сопровождающаяся зрительными галлюцинациями, чувством deja vu и deja pense («уже виденное» — фр.), при которых зрительный опыт и мысли имеют странные дружественные отношения, появляются автоматизмы, сопутствующие амнезии, компульсивное поведение, гнев и беспокойство.
У больных с односторонним повреждением первичной слуховой коры отмечается незначительная потеря слуха. При нарушении слухового узнавания (слуховая агнозия) поражены ассоциативные слуховые поля. Больные, как правило, отмечают, что они могут слышать звуки, но не могут их идентифицировать.
На вентральной поверхности височной доли обнаруживается область, участвующая в узнавании лица. Повреждение этой области (чаще при нарушении кровообращения в области ба-зилярной артерии мозга), что, как правило, распространяется и на смежную затылочную долю (поля 20, 21), обоих полушарий, сопровождается развитием прозопагнозии (неспособность узнавать лица). Вообще, это состояние не сопровождается другими неврологическими признаками. Больной может читать и называть объекты. В случаях, когда больной слышит голос знакомого человека, он может немедленно идентифицировать его.
Оставшаяся часть верхней височной извилины занята областью 22 (слуховая ассоциативная кора), которая получает большое количество афферентов от полей 41 я 42 к отдает волокна теменной и затылочной долям. Область 22 также вовлечена в функции языка. Повреждение верхней части 22-го поля приводит к возникновению трудностей в понимании языка.
Височные доли особенно важны в функции памяти. Например, стимуляция слуховой ассоциативной коры у больных во время нейрохирургических операций вызывает сложные воспоминания как слухового, так и зрительного характера. При обширных повреждениях височной доли возникает нарушение памяти.
Лимбическая доля(рис. 4.1.14).Лимбичес-кая система представляет комплекс образований конечного, промежуточного и среднего мозга. Фактически «лимбическая система» совпадает с ранее применявшимся понятием — «обонятельный мозг» [3, 4, 6, 8, 16, 273].
Анатомия головного мозга
 | |||
 | |||
|
| К мозжечку |
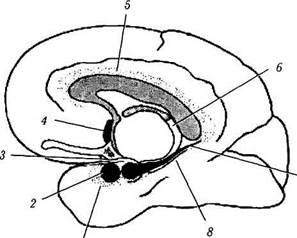
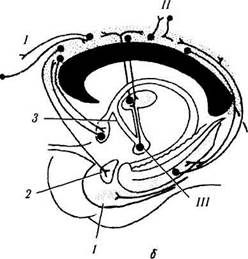
Рис. 4.1.14. Лимбическая система (по Brodsl, 1992):
а — расположение структур лимбической системы (/ — энтори-альное поле; 2 — миндалина; 3 — крючок; 4 — септальное ядро; 5 — поясная извилина; б — свод; 7 — гиппокамп; 8 — парагип-покампальная извилина); б — связи поясной извилины (/ — лобная ассоциативная кора; // — теменная и височная ассоциативная кора; /// — сосцевидное тело). Поясная извилина связана с ассоциативными полями коры головного мозга и лимбической системой, обеспечивая взаимодействие между ними
Если новая кора (неокортекс) управляет пространственно-временными взаимоотношениями организма с окружающей средой, а также отвечает за формально-логическое мышление и стереогностические способности, то лимбическая система обусловливает эмоциональные функции и побуждение к действию, а также процессы научения и памяти. Контролирует она и гомеостаз. Немаловажной ее функцией является организация поведенческих реакций индивидуума в ответ на воздействия внешней среды и изменение внутренней среды организма. Эти реакции направлены на сохранение особи и опосредуются через низшие центры промежуточного мозга.
Лимбическая доля имеет какое-то отношение и к функции обоняния, поскольку раздражение или повреждения области гиппокампа сопровождаются ощущением запахов.
Лимбическую долю образуют: подмозолис-тая область (area subcallosa), поясная извилина (gyrus cinguli), перешеек поясной извилины (isthmus gyri cinguli), парагиппокампальная извилина (gyrus parahippocampalis), гиппокамп (аммонов рог, зубчатая извилина и основание гиппокампа, или субикулум) и диагональная связка Брока. Кроме того, ряд исследователей относят к ней и филогенетически старые структуры обонятельного мозга (обонятельные луковицы, обонятельные бугорки и области коры, расположенные над миндалинами) (см. рис. 4.1.14).
К подкорковым структурам лимбической системы относятся миндалины (corpus amygda-loideum), септальные ядра и переднее талами-ческое ядро.
Афферентные и эфферентные связи структур лимбической системы как между собой, так и с другими отделами головного мозга чрезвычайно разнообразны. Наиболее выражены ре-ципрокные связи ее с гипоталамусом.
Гипоталамус и сосковидные (мамиллярные) тела соединяются с гиппокампом и септальной областью посредством свода, с миндалиной — посредством терминальной полоски и амигдало-фугального пучка, а с обонятельным мозгом — посредством медиального пучка переднего мозга. Через гипоталамус и сосцевидные тела лимбическая система соединена со средним мозгом.
Лимбическая система сообщается с новой корой в области лобной и височной долей. Височные области отвечают главным образом за передачу информации от зрительной, слуховой и соматической коры миндалине и гиппокампу. Лобные области служат основным отделом новой коры, регулирующим лимбические зоны.
Базальные ганглиипредставляют собой подкорковое скопление нейронов в виде группы ядер (рис. 4.1.15; 4.1.16, см. цв. вкл.). Контролируют они двигательную активность тела [3, 4, 6,8, 9, 140].Эта функция базальных ганглиев была установлена на основании клинических наблюдений. При их повреждении отмечается нарушение двигательной активности как в виде отсутствия возможности инициировать необходимые движения, так и отсутствия возможности подавлять движения.
При болезни Паркинсона, возникающей при повреждении базальных ганглиев, у больных лицо «невыразительное» [499]. Это связывают с нарушением двигательной активности мимических мышц и контроля движений глаза, в частности нарушения саккадических движений. Именно по этой причине мы более подробно рассмотрим нейроанатомию этой области.
Различают три основных скопления подкорковых ядер, которые называются полосатое
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ


|
| 15 |
| 16 |
| 17 |
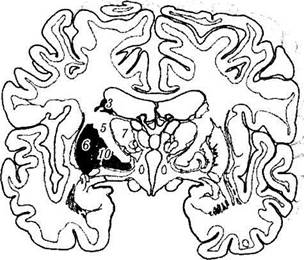
Рис. 4.1.15. Базальные ганглии:
а — схематическое изображение расположения структур, относящихся к базальным ганглиям; б — фронтальный срез головного мозга, проходящий через структуры, относящиеся к базальным ганглиям (/ — мозолистое тело; 2— рог бокового желудочка; 3—тело хвостатого ядра; 4—свод; 5—внутренняя капсула; 6 — островок; 7 — зрительный бугор; 8— скорлупа; 9 — ограда; 10— бледный шар; // — подбугорная область; 12 — безымянная субстанция; 13 — зрительный тракт; 14 — задняя нижняя часть хвостатого ядра; /5 — миндалина; 16 — височный рог бокового желудочка; 17 — лобный рог бокового желудочка; 18 — третий желудочек)
тело {corpus striatum), ограда (claustrum) и миндалевидное тело (corpus amigdaloideum).
1. Corpus striatum состоит из двух частей — хвостатого ядра (nucleus caudatus) и чечевице-образного ядра (nucleus lentiformis).
Хвостатое ядро лежит выше и медиальнее чечевицеобразного ядра, отделяясь от последнего прослойкой белого вещества, называемой внутренней капсулой (capsula interna). Утолщенная передняя часть хвостатого ядра, его головка (caput nuclei caudati) образуют латеральную стенку переднего рога бокового желудочка, задний же утонченный отдел хвостатого ядра (corpus et cauda nuclei caudati) тянется
назад по дну центральной части бокового желудочка. С медиальной стороны nucleus caudatus прилегает к зрительному бугру, отделяясь от него полоской белого вещества (stria termina-lis). Спереди и снизу головка хвостатого ядра доходит до переднего перфорированного вещества (substantia perforata anterior). В этом месте головка соединяется с чечевицеобразным ядром (с частью, называемой putamen). Кроме этого широкого соединения обоих ядер с вентральной стороны, имеются еще тонкие полоски серого вещества, располагающиеся вперемешку с белыми пучками внутренней капсулы. Они послужили причиной названия «полосатое тело» (corpus striatum).
Чечевицеобразное ядро (nucleus lentiformis) залегает латерально от хвостатого ядра и зрительного бугра, отделенное от них внутренней капсулой. На горизонтальном разрезе полушария медиальная поверхность чечевицеобразного ядра, обращенная к внутренней капсуле, имеет форму угла с верхушкой, направленной к середине. Передняя сторона угла параллельна хвостатому ядру, а задняя — зрительному бугру. Латеральная поверхность немного выпукла и обращена к латеральной стороне полушария в области островка. Спереди и вентрально, как было уже указано, чечевицеобразное ядро сливается с головкой хвостатого ядра.
На фронтальном разрезе чечевицеобразное ядро имеет форму клина, верхушка которого обращена в медиальную сторону, а основание — латерально. Чечевицеобразное ядро двумя параллельными белыми прослойками (laminae medullares) разделяется на три членика. Латерально расположенный участок темно-серого цвета называется скорлупой (putamen), а два медиальных, более светлых, вместе носят название бледного шара (globus pallidus).
Отличаясь уже по своему макроскопическому виду, бледный шар (globus pallidus) имеет также и гистологическую структуру, которая существенно отличается от строения других частей полосатого тела.
Ввиду всех этих особенностей бледный шар выделяют в особую морфологическую единицу под названием pallidum, тогда как обозначение striatum оставляют только за putamen и nucleus caudatus. Вследствие этого термин «чечевицеобразное ядро» теряет свое прежнее значение и может употребляться только в чисто топографическом смысле. При этом вместо прежнего названия corpus striatum хвостатое и чечевицеобразное ядра именуют стриопалли-дарной системой.
Стриопаллидарная система представляет собой главную часть экстрапирамидной системы, а кроме того, она является высшим регулирующим центром вегетативных функций в отношении теплорегуляции и углеводного обмена, доминирующим над подобными же вегетативными центрами в гипоталамусе.
Анатомия головного мозга
2. 
 Ограда (claustrum) представляет собой
Ограда (claustrum) представляет собой
тонкую пластинку серого вещества, заложен
ную в области островка, между ним и putamen.
От последнего она отделяется прослойкой бе
лого вещества, называемой наружной капсулой
(capsula externa).
3. Миндалевидное тело (corpus amygdaloi-
deum) расположено под putamen в переднем
конце височной доли. Миндалевидное тело, по-
видимому, относится к подкорковым обонятель
ным центрам и к лимбической системе. В нем
оканчивается идущий из обонятельной доли
и переднего перфорированного вещества (sub-
stantia perforata anterior) пучок волокон, отме
ченный при описании зрительного бугра под
названием stria terminates.
Роль миндалины у человека недостаточно изучена. В основном, роль миндалины выявлена на основании исследования неврологических больных. Благодаря этим исследованиям установлено, что механическое или электрическое раздражение миндалины у человека часто приводит к возникновению страха или других реакций эмоционального характера. Таким образом, миндалина у человека участвует в процессах эмоционального восприятия [136, 492]. Показано также, что при двухстороннем повреждении миндалин возникает потеря «эмоциональной» памяти, появление «испуганного» выражения лица, нарушение интеграции обонятельной и зрительной информации [327, 328].
Хорошо известно, что человек обладает способностью установления настроения и эмоционального состояния другого индивидуума [429]. В последнее время выявлено участие миндалины в этом сложном физиологическом процессе. При определении эмоционального состояния человека наш мозг улавливает и анализирует еле заметные изменения выражения лица, и в первую очередь направленность взора [36, 295, 312]. Невральные механизмы, лежащие в основе этих процессов, точно не ясны. Тем не менее установлено, что «точная» установка взгляда, психофизиологически оцениваемая как наличие «внимания», определяется верхней частью височной коры и миндалиной [331]. При исследовании больных аутизмом выявлено, что нередко у них выявляется аномалия миндалины или ее повреждение [128, 250]. Непосредственные механизмы, лежащие в основе социального суждения о других личностях на основании определения направленнности взгляда, остаются в значительной степени неизвестными.
К базальным ганглиям в последнее время относят также черную субстанцию (substantia nigra) и субталамические ядра. Причем в черной субстанции различают 2 части — ретикулярную [pars reticulata) и компактную (pars compacta) [3, 4, 6, 8, 9].
Морфологическими и электрофизиологическими методами показано, что большая часть афферентных сигналов, приходящих к базаль-
ным ганглиям, поступает в полосатое тело (см. рис. 4.1.16). Эти сигналы исходят из многих источников, основными из которых являются:
1) все области коры больших полушарий;
2) внутрипластинчатые ядра зрительного бугра
и 3) черная субстанция (по дофаминэргичес-
кому пути).
От внутреннего отдела бледного шара берет начало самый важный из всех эфферентных путей базальных ганглиев. Заканчивается этот путь преимущественно в зрительном бугре и крыше среднего мозга.
Таким образом, базальные ганглии играют роль промежуточного звена в цепи, связывающей двигательные области коры со всеми ее остальными областями. Основной их функцией является «планирование» двигательной активности.
В последние годы установлено, что базальные ганглии совместно с некоторыми корковыми областями также определяют некоторые познавательные функции, включая внимание, память [21, 246, 531]. Понятие «внимание», в широком смысле слова, представляет собой «процесс выбора» [245]. Учитавая то, что процесс ориентации в пространстве и выбора цели интереса, участие в котором принимают саккадические движения глаз, связан с базальными ганглиями, становится ясна значительная роль базального комплекса в процессах ориентации посредством саккад. При этом особое место в контроле сак-кад глаза играет черная субстанция, в частности ее ретикулярная часть. Как анатомически, так и электрофизиологически доказано наличие ее связи с верхними бугорками четверохолмия [18, 41, 161, 433, 585]. Основной функцией нейронов черной субстанции является торможение активности нейронов верхних бугороков четверохолмия, участвующих в генерации саккад.
В двигательной активности глаз принимает участие и хвостатое ядро, имеющее связи с черной субстанцией и наружными бугорками четверохолмия [96, 107, 201]. Возбуждение хвостатого ядра приводит к саккадическим движениям глаза в противоположную от возбуждаемого ядра сторону [310, 401].
Субталамическое ядро также участвует в движении глаз. На это ядро проецируются бледный шар [506], кора лобной области [99, 396]. От субталамического ядра отходят волокна к черной субстанции, наружному и внутреннему сегментам бледного шара [408].
Функция бледного шара в глазодвигательной активности менее ясна. Это связано с тем, что бледный шар имеет большое количество связей между различными ядрами базальных ганглиев и мало связей с нервными центрами, расположенными вне базальных ганглиев [96, 201, 235, 309, 521]. Глазодвигательные нейроны были обнаружены лишь в дорзальной части бледного шара [293], т. е. в части, принимающей входы от хвостатого ядра [235].
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 Роль базальных ганглиев в движении глаз подтверждается многочисленными клиническими наблюдениями. При болезни Паркинсона, сопровождающейся дегенерацией базальных ганглиев и серого вещества среднего мозга и прерывающей фронтомезенцефалические пути, развивается прогрессивный надъядерный паралич. У таких больных скорость саккадических движений глаза уменьшена. Саккады, как правило, гипометрические, что связано с нарушением функции «памяти саккады» [96, 201, 219, 244, 247, 569]. Часто движение глаз не может быть завершено в полном объеме. Вертикальные движения глаза повреждаются более значительно, чем горизонтальные. У больных также выявляется осевая ригидность и деменция. Близкие изменения выявлены и при болезни Хунтингтона [551].
Роль базальных ганглиев в движении глаз подтверждается многочисленными клиническими наблюдениями. При болезни Паркинсона, сопровождающейся дегенерацией базальных ганглиев и серого вещества среднего мозга и прерывающей фронтомезенцефалические пути, развивается прогрессивный надъядерный паралич. У таких больных скорость саккадических движений глаза уменьшена. Саккады, как правило, гипометрические, что связано с нарушением функции «памяти саккады» [96, 201, 219, 244, 247, 569]. Часто движение глаз не может быть завершено в полном объеме. Вертикальные движения глаза повреждаются более значительно, чем горизонтальные. У больных также выявляется осевая ригидность и деменция. Близкие изменения выявлены и при болезни Хунтингтона [551].
Мозолистое тело (corpus callosum). Мозолистое тело представляет собой большой пучок волокон (состоит примерно из 250 млн волокон), соединяющих между собой оба полушария (рис. 4.1.17). Основной функцией этого пучка является передача информации из одного полушария в другое [3, 4, 6, 8, 9].
Основными функциями мозолистого тела являются:
1. Корреляция изображений на правой и
левой половинах поля зрения.
2. Интеграция ощущений, поступающих от
парных конечностей, необходимая для обучения
двигательной координации.
3. Интеграция процессов внимания и акти
вации в полушариях.
У больных, у которых с терапевтической целью была произведена перерезка мозолистого тела, нарушений функций мозга в повседневной жизни не наблюдалось. Они выявлялись лишь с помощью специальных тестов. У других больных развиваются иногда довольно значительные нарушения поведения. Наиболее часто проявляется это «словесной слепотой», «словесной глухотой» и потерей координации движений правой и левой конечностей.
Дата добавления: 2015-03-26; просмотров: 1557;