Анатомия головного мозга. ножку мозга (pedunculus cerebri), а оттуда через мост — в продолговатый мозг.
 ножку мозга (pedunculus cerebri), а оттуда через мост — в продолговатый мозг.
ножку мозга (pedunculus cerebri), а оттуда через мост — в продолговатый мозг.
2. Корково-ядерный путь (tractus cortico-
nuclearis) представляет собой комплекс прово
дящих путей к двигательным ядрам черепно-
мозговых нервов. Начавшись от пирамидных
клеток коры нижней части предцентральной
извилины, они проходят через колено внутрен
ней капсулы и через ножку мозга, затем всту
пают в мост и, переходя на другую сторону,
оканчиваются в двигательных ядрах противопо
ложной стороны, образуя перекрест. Неболь
шая часть волокон оканчивается без перекрес
та. Так как все двигательные волокна собраны
на небольшом пространстве во внутренней кап
суле (колено и передние две трети задней нож
ки ее), то при повреждении их в этом месте
наблюдается односторонний паралич противо
положной стороны тела.
3. Корково-мостовой путь (tractus cortico-
pontini) идет от мозговой коры к ядрам моста.
Выделяются пути, направляющиеся от коры
лобной доли (tractus frontopontinus), затылоч
ной (tractus occipitopontinus), височной (tractus
temporopontinus) и теменной (tractus parieto-
pontinus). В качестве продолжения этих путей
из ядер моста идут волокна в мозжечок в со
ставе его средних ножек. При помощи этих
путей кора большого мозга оказывает тормоз
ное и регулирующее влияние на деятельность
мозжечка.
4. Таламо-кортикальные и корково-талами-
ческие волокна (fibrae thalamocorticalis et
corticotalamici) направляются от таламуса к
коре и обратно от коры к таламусу. Из воло
кон, идущих от таламуса, необходимо выде
лить так называемую центральную таламичес-
кую лучистость. Таламическая лучистость явля
ется конечной частью чувствительного пути,
направляющегося к центру кожного чувства в
постцентральную извилину. Выходя из. лате
ральных ядер таламуса, волокна этого пути
проходят через заднюю ножку внутренней
капсулы позади пирамидного пути. Это место
названо чувствительным перекрестом, так как
здесь проходят и другие чувствительные пути,
а именно: зрительная лучистость (radiacio opti-
са) и слуховая лучистость (radiacio acustica).
Цитоархитектоника.Кора мозга довольно строго организована как в горизонтальной, так и вертикальной плоскостях. Горизонтальная организация сводится к тому, что существует иерархия распределения различных типов нейронов в виде слоев. Вертикальная организация основана на существовании определенной вертикальной ориентации отростков нейронов различных типов.
Слоистость. На всем протяжении кора состоит из слоев тел клеток. Учитывая количество слоев в различных участках, кору можно разделить на два основных типа: неокортекс и аллокортекс. Неокортекс состоит из 6 слоев
нервных клеток, а в аллокортексе их меньше. Специфическим типом аллокортекса является архикортекс, который состоит из трех клеточных слоев. У человека к архикортексу относится гиппокамп. Большая часть коры (90%) относится к неокортексу.
Увеличение площади коры головного мозга в филогенезе происходило путем образования извилин и борозд. Фактически две трети коры мозга человека погружены в толще тканей в пределах этих извилин.
|
Как вертикальная, так и тангенциальная организация неокортекса отличается особенностями строения составляющих ее нейронов. Морфологических типов нейронов насчитывается более 60. Различают следующие два основных типа клеток — пирамидные и непирамидные (рис. 4.1.6). Пирамидные клетки по разным оценкам составляют от 50 до 80% всех нейронов коры. Пирамидные клетки характеризуются треугольной формой тела. От их апикальной поверхности отходит длинный покрытый шипи-ками дендрит, направляющийся в молекулярный слой коры, где он и ветвится. От базаль-ной и латеральной частей тела нейрона отходит 5—16 коротких дендритов, которые ветвятся в том же слое, где располагается тело нейрона.
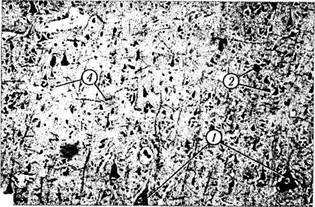
Рис. 4.1.6.Цитологические особенности нейронов коры головного мозга:
а — окрашивание по Нисслю; б, в — импрегнация серебром
(/ — пирамидные; 2 — корзинкоподобные; 3 — звездчатые; 4—
глиальные клетки)
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 От середины базальнои поверхности тела отходит аксон, направляющийся в белое вещество. На расстоянии 60—90 мкм от тела клетки аксон начинает разветвляться.
От середины базальнои поверхности тела отходит аксон, направляющийся в белое вещество. На расстоянии 60—90 мкм от тела клетки аксон начинает разветвляться.
Размер пирамидных клеток колеблется от 10 до 50 мкм. Гигантские пирамидные клетки (клетки Беца) имеют размер 100 мкм и обнаруживаются в пятом слое 4-го и 6-го полей коры. Основной функцией пирамидных клеток является интеграция поступающей информации внутри коры и образование эфферентных путей.
Клетки, не относящиеся к пирамидным нейронам, подразделяются на несколько типов в зависимости от их морфологии. Наиболее распространенными являются звездоподобные, кор-зинчатые, аксо-аксонные клетки, клетки «канделябры», клетки с двойным «букетом» дендри-тов, веретенообразные, горизонтальные (клетки Кахала) и клетки Мартинотти (рис. 4.1.6, 4.1.7). Основной функцией непирамидных клеток является интеграция нейронных цепей внутри коры.
Слои неокортекса (рис. 4.1.8). Как указано выше, кора головного мозга обладает послойной организацией нейронов и их отростков [4, 6—9, 11, 76, 94, 397, 580]. Большинство авторов выделяют шесть слоев:
I. Молекулярный (плексиформный) слой
(молекулярная пластинка; lamina molecularis
(plexiformis)).
Этот слой наиболее поверхностный (расположен под мягкой мозговой оболочкой). Состоит он из плотно переплетенных аксонов и дендритов. Источником волокон являются нейроны коры головного мозга (главным образом, звездчатые клетки), пирамидные клетки (расположены в более глубоких слоях). В этом слое располагаются и терминалы афферентных волокон из других отделов центральной нервной системы. Содержит он сравнительно небольшое количество горизонтальных клеток Кахала с длинными ветвящимися дендритами. Их аксоны участвуют в образовании тангенциального сплетения волокон этого слоя.
II. Наружный зернистый слой (наружная зер
нистая пластинка; lamina granularis externa).
Этот слой лежит кнутри от плексиформного слоя. Он называется зернистым из-за большого скопления ядер клеток. Состоит этот слой из тел нейронов, их дендритов и аксонов. Часть клеток пирамидной формы, некоторые звездчатые (мультиполярные). В этом слое аксоны зрительного нерва и дендриты входят в контакт с дендритами нейронов коры. Афферентные волокна формируют многочисленные синапсы с нейронами коры, особенно с апикальными дендритами пирамидных клеток. Дендриты нейронов ветвятся и поднимаются в молекулярный слой, а аксоны либо уходят в белое вещество, либо образуют дуги и также направляются в молекулярный слой.
III. Слой пирамидных клеток (наружная пира
мидная пластинка; lamina pyramidalis externa).
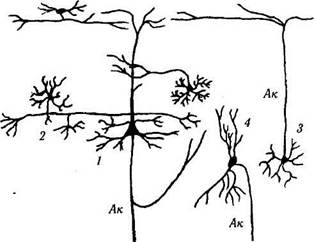
Рис. 4.1.7. Некоторые типы нейронов коры большого мозга:
/ — пирамидный нейрон; 2 — корзинкоподобный нейрон; 3 — звездчатый нейрон; 4 — веретенообразный нейрон. Ак — аксон

Рис. 4.1.8. Схематическое изображение пяти основных
типов послойной организации коры головного мозга
(по Williams et al., 1995):
а—двигательная кора. Отмечается отсутствие гранулярных слоев и наличие скоплений гигантских пирамидных клеток (Беца); б — премоторная кора. Отсутствуют гранулярные слои и гигантские пирамидные клетки. Крупные пирамидные клетки обнаруживаются в V слое; в — сенсорная кора (постцентральное поле). Хорошо выражен гранулярный слой; г — зрительная чувствительная кора. Редукция III слоя и существенное утолщение IV слоя. Последний пересекается зрительной полоской; д— зрительная «психическая» кора. Гранулярный слой хорошо развит, но крупные клетки в V слое отсутствуют
Дата добавления: 2015-03-26; просмотров: 1238;