Механизмы формирования временной связи. Память
Локализация временных связей. Физиологические основы механизмов образования временных связей. Основу пластичных взаимоотношений между организмом и окружающей средой составляют условные рефлексы. Физиологическую основу условных рефлексов составляет процесс замыкания временной связи.
Временная связь - это совокупность нейрофизиологических, биохимических и ультраструктурных изменений мозга, возникающая в процессе сочетания условного и безусловного раздражителей и формирующая определенные взаимоотношения между различными мозговыми образованиями. Механизм памяти фиксирует эти взаимоотношения, обеспечивая их удержание и воспроизведение.
Исходной предпосылкой замыкания временной связи является общефизиологический феномен повторения пути - усиление рефлекторного ответа организма при одновременном или последовательном раздражении одного и того же рецептивного поля. То есть одно раздражение своими следовыми эффектами облегчает появление ответа на действие следующего. Этот процесс непосредственно связан с явлением суммации возбуждения (И.М. Сеченов). Оно заключается в возникновении действенного центра возбуждения в нервном центре в условиях применения ряда последовательных раздражения, каждое из которых само по себе не способно вызвать какой-либо ответ.
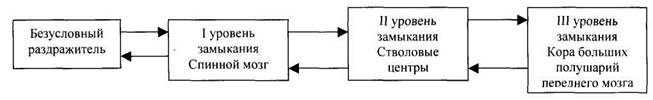
И.П. Павлов считал, что замыкание временных связей происходит в коре больших полушарий переднего мозга между тем пунктом, который воспринимает раздражитель, и корковым представительством безусловного рефлекса.
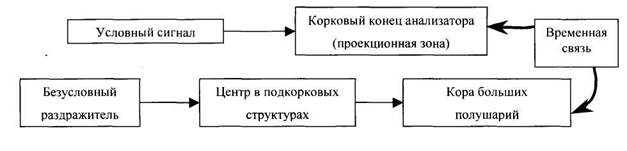
Э.А. Асратян пришел к выводу, что центральная часть дуги безусловного рефлекса не однолинейна, а имеет многоуровневую структуру, то есть состоит из многих ветвей, которые проходят различные уровни центральной нервной системы, наивысшая часть дуги проходит через кору большого мозга, являясь корковым представительством данного безусловного рефлекса и осуществляет кортиколизацию соответствующих функций.
Если сигнальный и подкрепляющий раздражители вызывают собственно безусловные рефлексы, то они и составляют нейросубстрат условного рефлекса.
Таким образом, образование условного рефлекса - это синтез двух (и более) различных безусловных рефлексов.
В процессе формирования условных рефлексов в корковых проекциях сигнального и подкрепляющего раздражителей происходит функциональная перестройка. Постепенно сигнальный стимул начинает вызывать несвойственную ему ранее условную реакцию, и в то же время изменяется его «собственная» безусловно-рефлекторная реакция. Благодаря этому соответствующее ему эффекторное выражение условного раздражителя в процессе обучения угасает (за счет процессов внутреннего торможения). В то же время в эфферентной части дуги подкрепляющего раздражителя возбудимость возрастает и условный стимул становится эффективным для запуска несвойственной ему ранее эффекторной реакции.
При действии неспецифического (индифферентного) раздражителя, вызывающего определенные безусловные рефлексы, образуется условный рефлекс с двусторонней временной связью, наличие которой является общей закономерностью «переключения» в ВНД. Прямые и обратные условные связи могут быть близки по силе, стойкости, возбудимости и другим свойствам, но могут быть также значительно отличаться друг от друга по этим показателям.
Принцип переключения имеет место не только в деятельности коры головного мозга, но именно здесь он достигает высшего уровня развития и делает деятельность лабильной и разнообразной. В «переключательной» функции коры участвуют тонические (настроечные, перестроечные) и фазические (завершающие, командные, исполнительные) механизмы. Настроечный механизм регулирует общий тонус, создает определенный уровень возбудимости и работоспособности нервных центров. Фазический механизм запускает процессы, которые осуществляют ту или иную реакцию. Все эти свойства коры больших полушарий обеспечивают ей способность контролировать и регулировать функциональное состояние мозга, а также свое собственное в зависимости от внешней и внутренней стимульной программы.
Регуляция деятельности нервных центров при изменении условий происходит по типу укороченных условных рефлексов, то есть тонических рефлексов, не имеющих обычного завершающего эффекторного конца, а создают лишь определенное состояние нервных центров в процессе условно-рефлекторной деятельности.
Доминанта. Доминанта — это господствующий очаг возбуждения, предопределяющий характер текущих реакций нервных центров в данный
момент. Такой центр может возникнуть на разных «этажах» центральной нервной системы при достаточно длительном действии гуморальных и рефлекторных раздражителей и характеризуется следующими параметрами:
- наличие повышенной возбудимости;
- инерционность, обусловленная длительными следовыми процессами;
- способность к суммации и сопряженному торможению других центров, функционально несовместимых с деятельностью центров доминантного очага.
В целом доминанта определяет вероятность возникновения той или иной рефлекторной реакции в ответ на текущее раздражение, характеризуется направленностью и создает определенный вектор поведения. Доминантный очаг возбуждения может возникнуть не только за счет гуморальных воздействий, но и под влиянием нервных сигналов, изменяющих возбудимость центральных нейронов.
Рассматривая доминанту как общий принцип работы нервных центров, Ухтомский считал, что она связана с возбуждением целого ряда нервных центров (их констелляция), которые временно кооперируются при выполнении биологически важной функции. Это объединение дает динамический функциональный орган, сообщающий в данный момент организму единство действия. Наряду с этим при в констелляции нервных центров отдельные ее компоненты могут участвовать в выполнении других рефлекторных актов.
Постепенно формируясь, рабочая констелляция проходит несколько стадий.
1. Стадия укрепления наличной доминанты по преимуществу. На этой стадии происходит образование доминанты под влиянием конкретных раздражителей. При этом в констелляцию вовлекаются не только необходимые для этого акта нервные центры, но и посторонние клеточные группы. На данной стадии наблюдается диффузная отзывчивость доминирующей констелляции к различным раздражениям. Постепенно происходит конкретизация раздражения, на который формируется определенный ответ.
2.Стадия выработки адекватного раздражителя для данной доминанты. 3.Стадия выделения данного комплекса раздражителей из среды. На этих стадиях происходит подбор биологически интересных рецепций для доминанты, что приводит к образованию. Новых адекватных поводов для той же доминанты. Результатом этих изменений является более экономичное выполнение реализации доминанты за счет затормаживания ненужных в данный момент процессов. Приповторении доминанты повторяется и свойственный только ей единыйритм возбуждения.
После прекращения существования данной доминанты в коре больших полушарий остается так называемый эскиз, позволяющий наиболее экономично восстановить в случае необходимости структуру доминирующего очага. Это в свою очередь обеспечивает более быстрое образование поведенческого акта в случае действия «знакомых» раздражителей. Более того, на базе имеющегося доминантного эскиза возможна достаточно быстрая перестройка констелляции нервных центров при подборе наиболее интересных для доминанты раздражений из новой среды.
Нейронные механизмы обучения. Выделяют две основные гипотезы обучения - синаптическую и мембранную. Синаптическая гипотеза заключается в том, что механизм формирования условных рефлексов представляет собой изменение эффективности синапсов. Мембранная гипотеза утверждает, что основным механизмом условного рефлекса является изменение свойств возбудимой постсинаптической мембраны.
Нервные механизмы обучения обеспечиваются благодаря двум основным свойствам нервной системы - реактивности и пластичности.
Реактивность - это способность отвечать на раздражения, пластичность -способность изменять реактивность под влиянием последовательных раздражений. В свою очередь реактивность определяется возбудимостью постсинаптической мембраны и эффективностью соответствующих синапсов.
В процессе реактивности нейронов при образовании условных рефлексов значение имеют механизмы привыкания (пресинаптической депрессии медиатора) и сенситизации, или сенсибилизации
(посттетанической потенциации). Привыкание нейронов заключается в снижении вероятности ответа на каждый последующий стимул в серии. Сенситизация - усиление реакции при повторных применениях стимула.
Сенсибилизация в реагировании нейронов коры возникает в том случае, если раздражители прямо или косвенно связаны с мотивационной сферой животного или человека.
Привыкание ответов тех же нейронов наблюдается в тех случаях, когда исследуемые раздражители были индифферентными или хорошо знакомыми животному.
Пластичность нервных процессов обеспечивается суммацией рефлексов, в основе которой лежат сенсибилизация и фасилитация (облегчение запуска именно данной ответной реакции).
Те структуры, где изменения происходят в первую очередь, рассматривают как наиболее существенные для обучения. Критерием вовлеченности нейрона в условно-рефлекторный процесс является характер предшествующей реакции, из чего вытекает необходимость существования «критических синапсов», в которых происходят прежде всего изменения, обусловливающие распространение импульса по новому пути.
В качестве наиболее значимых для обучения отделов мозга отмечены фронтальная и сенсорная кора, а также гиппокамп.
Торможение условных рефлексов. Виды торможения. Успех индивидуальной адаптации организма к разнообразным
условиям его жизни, вся динамика условно-рефлекторной деятельности зависят от сложных взаимоотношений между возбудимыми и тормозными процессами в головном мозге.
Процессы торможения в центральной нервной системе открыл и подробно описал И.М. Сеченов (1861). Он рассматривал центральное торможение как деятельность (возбуждение) специализированных тормозных систем по аналогии с хорошо изученным к тому времени тормозным эффектам блуждающего нерва на сердце.
Ч. Шеррингтон (1906) развивал представление об активном торможении мышц-антагонистов в процессе рефлекторного акта.
А.А. Ухтомский (1923), опираясь на предшествующие взгляды, выдвинул представление о сопряженном торможении при формировании доминанты.
И.П. Павлов, изучая условные рефлексы и их взаимоотношения, наблюдал торможение (угнетение) условных рефлексов при действии посторонних или сильных раздражителей, а также слабых - при болезненном состоянии организма. Он считал, что баланс между возбуждением и торможением определяет внешнее проявление поведения животных и человека, и выдвинул собственную схему классификации видов торможения при условно-рефлекторной деятельности.
Внешнее (безусловное) торможение - это срочное подавление текущей условно-рефлекторной деятельности при действии посторонних для нее раздражителей, вызывающих какой-либо безусловный рефлекс. Данное
торможение не изменяет структуру условного рефлекса. Это врожденный тип (индукционное, по И.П. Павлову; сопряженное, по А.А. Ухтомскому).
Гаснущий тормоз - кратковременное выключение нервной деятельности, вызываемое незначительным раздражителем (в тихой аудитории пролетает муха);
Негаснущий тормоз - длительное отвлечение от активной деятельности (зубная боль, голод);
Запредельное (охранительное) торможение - возникает при действии раздражителя очень высокой мощности. «Охраняет» клетки головного мозга от избыточного расходования энергии. Зависит от функционального состояния нервной системы, возраста, эмоционального фона. Предел выносливости клетки в отношении раздражителей разной интенсивности называется пределом ее работоспособности. Чем выше этот предел, тем легче клетка переносит действие сверхсильных раздражителей. Крайним случаем запредельного торможения является оцепенение, возникающее у животных и человека под влиянием раздражителя очень большой мощности. Человек может впасть в состояние ступора - полной неподвижности.
Внутреннее (условное) торможение наблюдается при прекращении безусловного подкрепления условного рефлекса. Это торможение развивается медленно и изменяет структуру самого рефлекса.
Основными чертами условного торможения являются:
- развивается при неподкреплении раздражителей, которые постепенно приобретают свойства условного тормоза или отрицательного сигнала;
- поддается тренировке. Заторможенный рефлекс может самопроизвольно растормаживаться;
- способность к разным проявлениям условного торможения зависит от индивидуальных особенностей нервной системы: возбудимые хуже тормозятся;
- зависит от физиологической силы безусловного рефлекса, подкрепляющего условный сигнал;
- зависит от прочности ранее выработанного рефлекса;
- условное торможение может взаимодействовать с безусловным. В этих
случаях формируется так называемое растормаживание и (иногда)
суммация общего эффекта.
Виды условного торможения:
Угасательное развивается при отсутствии подкрепления условного сигнала безусловным.
Дифференцировочное торможение развивается при неподкреплении раздражителей, близких по свойствам к подкрепляемому.
Свойства дифференцировочного торможения:
- чем ближе дифференцируемые раздражители, тем труднее выработать на них дифференцировочное торможение;
степень торможения определяется силой возбуждения, развиваемого положительного условного рефлекса;
выработка этого торможения происходит волнообразно;
дифференцировочное торможение тренируемо.
Условный тормоз - меняются условия, меняется ответ.
Запаздывательное торможение - подкрепление отодвигается от начала действия условного раздражителя (кошка —» запах пищи —> мышь в зубах —► слюноотделение). В данном виде торможения выделяют недеятельную фазу (основное время действия условного раздражителя не подкрепляется безусловным рефлексом) и деятельную фазу (реакция на раздражитель).
К особым видам торможения относят сон, гипноз, наркоз.
Основные свойства нервных процессов. В основе всей условно-рефлекторной деятельности лежит неразрывное единство возбуждения и торможения. Оба нервных процесса участвуют в
формировании условного рефлекса, сложным образом взаимодействуя между собой. К основным свойствам нервных процессов относят:
1) Иррадиация (распространение). Нервный процесс, возникнув в одном центре, может распространяться и на другие участки. Чем интенсивнее нервный процесс, тем шире иррадиация. Иррадирующее торможение оказывает большее воздействие на клетки анализатора, чем ближе они находятся к тормозному очагу. Иррадиация возбуждения в коре головного мозга происходит гораздо быстрее, чем иррадиация торможения.
2) После широкой иррадиации наступает концентрация нервных процессов в месте своего возникновения. При своем концентрировании нервный процесс проходит в обратной последовательности все те пункты полей анализаторов, который он захватывал в своем поступательном движении.
3) Индукция нервных процессов - это свойство каждого из нервных процессов вызывать вокруг себя и после себя противоположный процесс. То есть имеющийся в коре головного мозга процесс возбуждения оказывает тормозящее действие на другие.
Дата добавления: 2015-03-14; просмотров: 5039;