Гликолиз. Р-ции. Регуляция. 3 страница
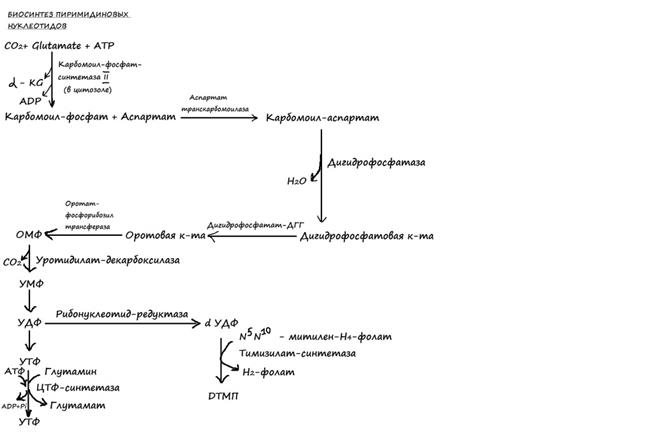
85. Биосинтез декарбоксирибонуклеотидов. Дезоксирибонуклеотиды обр-ся из рибонуклеотидов за счет восст-я остатка рибозы. донором протонов и электронов служит низкомолекулярный белок тироредуксин. Ингибиторы синтеза дезоксирибонуклеотидов тормозят деление клетки. Дезоксирибонуклеотиды образ-ся из рибонеклеотидов, за счет восстановл-я остатка рибозы: донором протонов и электронов служит тиоредоксин (содержит в SH группе), после окисления вновь восст-ся при помощи НАДФН(Н+). Общий путь синтеза: ХДФ-> дХДФ-> дХТФ. Исключение дезокси ТТФ, который образ-ся из УДФ. УДФ-> дезокси УДФ-> дезокси УТФ-> дезокси УМФ-> дезокси ТНФ-> дезокси ТДФ-> дезокси ТТФ. Синтез дезоксирибонукл-в в покоящих клетках минимален. Его активация набл-ся на стадиях, предшествующих делению кл.ингибиторы синтеза дезоксирибонукл-в тормозят репликацию и деление клетки.
86. Биосинтез тимидиловых нуклеотидов. Тимидиловые нуклеотиды входят в состав ДНК, содержащей дезоксирибозу. При помощи метода меченых атомов было показано, что этот синтез начинается не со свободнойдезоксирибозы, а путем прямого восстановления рибонуклеотидов у 2-го атома углерода. Химический смысл превращения рибонуклеотидов в дезоксирибо-нуклеотиды сводится к элементарному акту – восстановлению рибозы в 2-дезоксирибозу, требующему наличия двух атомов водорода. Непосредственным источником оказался восстановленный термостабильный белок тиоредоксин, содержащий две свободные SH-группы. Тиоредоксин легко окисляется, превращаясь в дисульфидную S-S-форму.
87.Пути регуляция активности ферментов.Метаболическая регуляция. Регуляция активности ферментов-ферменты являются регулируемыми катализаторами. В качестве регуляторов могут выступать метаболиты, яды. Различают:- активаторы – вещества, увеличивающие скорость реакции; - ингибиторы – вещества, уменьшающие скорость реакции. Активация ферментов. Различные активаторы могут связываться либо с активным центром фермента, либо вне его. К группе активаторов, влияющих на активный центр, относятся: ионы металла, коферменты, сами субстраты. Активация с помощью металлов протекает по различным механизмам: металл входит в состав каталитического участка активного центра; металл с субстратом образуют комплекс; за счет металла образуется мосты между субстратом и активным центром фермента. Субстраты также являются активаторами. При увеличении концентрации субстрата скорость реакции повышается. По достижению концентрации насыщения субстрата эта скорость не изменяется. Синтез ферментов, контролирующих скорость метаболических путей, может изменяться под действием гормонов. Поскольку в этом случае происходит синтез новых белковых молекул, изменение активности происходит сравнительно медленно и чаще всего в ответ на изменение количества и состава поступающей пищи. Гормоны могут действовать как индукторы или репрессоры синтеза мРНК в ядре или как стимуляторы стадии трансляции белкового синтеза на уровне рибосом.
88. Гормональная регуляция метаболизма. Понятия о гормонах, их биологическое значение. Классификация гормонов. Для нормального функционирования многоклеточного организма необходима взаимосвязь между отдельными клетками, тканями и органами. Эту взаимосвязь осуществляют 4 основные системы регуляции: ЦНС и ПНС, эндокринная и паракринная системы, иммунная система через белки. Регуляторными механизмами метаболизма в разных органах, являются гормоны, а также низкомолекулярные белковые соединения – цитокины- они синтезируются в процессе иммунитета, действуют как факторы роста.Все гормоны классифицируют по химическому строению, биологическим функциям и механизму действия. По хим. строению гормоны делят на 3 группы:пептидные-белковые (адренокортикотропный, соматотропонин, тиреотропный, окситоцин), стероидные (альдостерон, кортизол, прогестерон) и непептидные (адреналин, норадреналин, тироксин).По биологич-м функциям. Инсулин, кортизол, соматотропонин- регулируют обмен углеводов,аминокис-т. Альдостерон- водно-солевой обмен. Тропные гормоны- синтез щитовидки.
89. Роль гормонов в обеспечении межклеточной сигнализации. Трансмембранная передача сигналов в клетке. Мембранные и внутриклеточные рецепторы. Сигнал передается ч/з жидкие среды организма (кровь, лимфу, межклеточную жидкость). Это гуморальная система связи. Иначе она называется эндокринная сигнализация. Эндокринные клетки выделяют гормоны. Гормон - Это химические вещ-ва, служащие сигналами для других клеток, расположенных на расстоянии. При получении сигнала клетки изменяют режим функционирования. Эндокринные клетки обычно собраны в специальные железы и выделяют гормоны во внеклеточную жид-ть, окружающую все клетки в тканях. Гормоны с кровотоком разносятся по всему телу. Реакция на него будет только тогда, когда он будет захвачен рецептором клетки, специфически для этого предназначенным. При его приеме происходит запуск определенного специфического процесса, который и приводит к корректировке работы организма. Работа эндокринных желез построена по системе каскада. Центральный сигнал приводит к запуску процессов через систему эндокринных желез. Гормоны в крови и в межклеточной жид-ти очень сильно разбавляются и способны действовать в чрезвычайно низких концентрациях. Несмотря на широкий спектр гормонов, продуцируемых эндокринной системой, все рецепторы на основании их локализации можно разделить па две обширные группы: 1) мембранные или поверхностные и 2) внутриклеточные рецепторы. Мембранные рецепторы располагаются в плазматической мембране клетки-мишени и взаимодействуют с белково-пептидными гормонами и катехоламинами. Внутриклеточные рецепторы находятся внутри клетки и взаимодействуют со стероидными и тиреоидпыми гормонами, которые представляют собой небольшие липофильные молекулы, с легкостью проникающие в клетку через плазматическую мембрану.
90. Механизмы действия пептидных гормонов. Роль и виды вторичных посредников.Пептидные гормоны – это водораствор. гормоны, вырабатываемые железами внутренней секреции для контроля различных функций организма. Кортиколиберин- в гипоталамусе, кортикотропин- в коре надпочечников. Соматостатин - в поджелудоч. железе и гипоталамусе. Инсулин- в поджелудоч. железе, участв в регуляции уровня сахара в крови. Тиреотропонин- в перед доле гипофиза, в щитовидке, повышает йод. Эритропоэтин- в почках, в печени, стимулир. образование эритроцитов. Выделяют 3 вида вторич. посредников: 1.циклические нуклеотиды- Цамф- участвует в действии кортикотропонина, дофамина, адреналина, гистамина, глюкагона. Повыш. содержание стимулирует распад молекул, усиливает секрецию HCL в желудке. 2. Метаболиты фосфатидилинозитола- связаны фосфолипазой С. Сущ-ет два вторич. посредника- инозитол-1,4,5-трифосфат и 1,2-диацилглицерол-активирует протеинкиназу С- ее длительная активация нарушает рост и деление клеток. 3.Ионы Ca- связывается с белком кальмодулином- активирует мембранную АТФазу, которая откачив. кальций из цитозоля.
91. Структура, ф-ции и мех-зм действия стероидных гормонов.Исх. соединением явл ацитил КоА. 1 этап: синтез мевалоновой к-ты. Ацитил КоА –> ацитоацитил КоА -> В-окси-В-митилглутарил КоА -> меволоновая к-та. Синтез мевалоновой к-ты происх.в цитоплазм. Она явл предш-ком для синтеза изопреноидов, витаминов А и К, убихинона, фитола. 2 этап: образ-е активного изопрена и его последовательная конденсация в сквален . При этом происх.замыкание в циклическую структуру типа циклопентанпергидрофенантрен. В этих реакциях затрачивается НАД(Н+), ф-ция происходит в цитоплазме без участия О2. 3 этап: превращение сквалена в холестерин. В ходе этих р-ций происх.дальнейшая циклизация в стероидную структуру и отщепление СО2. Р-ции идут на поверх-ти ЭПР. Образовавшийся холестерин может транспортир-ся в митохондрии и там превращ-ся в первичную и вторичную желчн.к-ты (фолиевую, холановую, хенодезоксихолевую, гликохолевую и таурохолевую). 4 этап: синтез стероидн.гормонов происх.в ЭПР. Холестерин превр-ся в прогестерон, а прогестерон синтезир-ся в глюкокартикоиды и минералкортикоиды. Муж. Половые гормоны синтезир-ся из прогестерона под действием индивидуальных гидроксилаз, из тестостерона синтезир-ся эстрон и вся группа жен.половых гормонов. Катаболизм гормонов. Белково-пептидные гормоны подвергаются протеолизу, распадаются до отдельных аминок-т. Эти аминок-ты вступают в р-ции дезаминирования, декарбоксилирования, трансаминирования и распадаются до конечных продуктов: NH3, CO2 и Н2О.
92. Гормоны гипоталамуса: Кортиколиберин (стимулир. образование трофина и кортикотрофина), тириолиберин (стимулирует секрецию пролактина и териотропина), соматолиберин (ингибирует секрецию гормона роста и териотропина), пролактостатин (ингибирует выделение пролактина), меланолиберин (стимулир.секрецию меланоцит-стимулирующего гормона), гонадолиберин (стимулир-т образование лютропина и фоллитропина), пролактолиберин (способствует выделению пролактина), меланостатин (ингибир-т секрецию меланоцит-стимулирующего гормона). Нейроны гипоталамуса синтез-ют гормоны и секретир-т их в капиллярн.сеть, доставляющую гормоны к клеткам гипофиза, где они в свою очередь тормозят или стимулир-т синтез тропных гормонов. Либерины действуют на кл.гипофиза с 7-ТМС рецепторами с реализацией аденилатциклазной системы или инозитолтрифосфатного мех-зма.
93. Гормоны гипофиза. Гормоны аденогипофиза: соматотропин (стимулир-т рост орг-ма, синтез белка в клетках, образ-е глюкозы и распад липидов), пролактин (регулир-т лактацию млекопит-х, инстинкт выхаживания потомства, дифференцировка различных тканей), тириотропин ( регулир-т биосинтез и секрецию гормонов щ/ж-зы), кортикотропин (регулир-т секрецию коры надпочечников), фолликуло- стимулирующий гормон (оказывает действие на фолликулы и кл.Лейдига, действию лютоинизирующего гормона, стимулир-т сперматогенез), лютоинизирующий гормон (регулир-т синтез жен.и муж.половых гормонов, стимулир-т рост и созревание фолликулов, овуляцию, образ-е и функционирование желтого тела яичника). Гормоны задн.доли гипофиза: окситоцин и вазопрессин принято считать гормонами задн.доли, хотя они образ-ся и секретир-ся нейронами гипоталамуса. Вазопрессин обеспечив-т сохранение воды в организме, стимулируя ее реабсобцию почечных канальцев. Окситоцин увелич-т сокращ-е матки во время родов, стимулир-т лактацию, регулирующую поведенческую активность связанную с беременностью, лактацией и уходом за потомством. Главными стимулами в освобождении окситоцина явл эстрогены и нервные импульсы, возникающие при раздражении грудных сосков. Прогестерон ингибир-т продукцию окситоцина.юУ мужщин окситоцин стимулир-т семявыносящие протоки, влияет на половую активность. Недостаточность вазопрессина – удельный вес мочи низкий.т.к. большое кол-во глюкозы не выдел-ся с мочой.
Регуляция водно-солевого обмена . Нарушение ВСО.В его регуляции приним.участие такие гормоны как: антидиуретический (вазопрессин), альдостерон,атрионатрийуретический,ренин-ангеотензивная система.При регуляции используются 2 принципа:1.регуляция осмотического давления 2.регуляция объема жидкости.Вазопрессин вырабатывается в гипоталамусе. Воздействия:1.увеличение реабсорбции воды в канальц.аппарате 2.вызывает сужение сосудов,особенно артериол и капилляров 3.уменьшает диурез и вызывает жажду. Вазопрессин-пептидный гормон,второй посредник циклический АМФ,он снижает осмотическое давление,но оно же влияет на реабсорбцию Na и Cl.Сигналом для синтеза явл повышен. осмотич. давление, при этом вырабатыв.рецепторы в гипоталамусе,где и синтез-ся вазопрессин.Далее он перемещается в заднюю долю гипофиза,где запас-ся в виде гранул и синтезируется по мере надобности.Атрионатрий уретический гормон образуется в клетках Пуркинье сердечой мышцы.Он действует подобно вазопрессину,способствует выведению Na,изменении клубочковой фильтрации.Нарушения водно-солевого обмена (ВСО)Они бывают: 1.Нарушения баланса (не соответствие м/д поступл-ем и вывед-ем), 2.Нарушения распределения (м\д внеклет-ым и внутрикл-ым пространством). В зависимости от содержания жид. в орг-ме и осматич. давления плазмы (ОДП) различают 6 состояний связанных с увеличением кол-ва внекл. жид-ти (гипергидратация) и его уменьшением (дегидратация): 1. Гипертонич. дегидратация: абсолютный или преобладающий дефицит жид-ти с повышением ОДП (потеря жид-ти, пневмония); 2. Гипотонич. дегидратация: дефицит воды и растворенных в ней в-в с понижением ОДП (хронич. пиелонефрит, прием дистил. воды); 3. Изотонич. дегидратация: дефицит воды и растворенных в ней в-в при норм. ОДП (рвота, диарея, кишечн. непроходимость, кровопотеря, ожоги, прием диуретиков); 4. Гипертонич. гипергидратация: избыток воды и растворенных в-в с повышением ОДП, клетки обезвоживаются с потерей калия (причины: параэнтеральное введение изотонич. или гипертонич. р-ров при недостаточности ф-ий почек, питье морской воды); 5. Изотонич. гепергидратация: избыток воды и растворенных в-в при норм. ОДП, появл-ся переферич. отеки, происходит задержка Na, гиперальдостраниум (причины: чрезмерное введение р-ров при заболев-х почек, цирроз печени с асцитом); 6. Гипотонич. гипергидратация: пересыщение водой с понижением ОДП (причины: чрезмерное введение ж-ти или безсолевых р-ов сахаров, безсолевая диета, прим-е диуретиков. Нарушения могут быть вызваны гормональным дисбалансом, наруш-м всасывания ионов в ЖКТ, наруш-я соотношений фильтрации реобсорбции и секреции ионов в почках.
95. Гормональная регуляция мочеобразования. На работу почек оказывают влияние многие гормоны, главными из которых являются антидиуретический гормон (АДГ), или вазопрессин, и альдостерон. Антидиуретический гормон (АДГ), или вазопрессин, способствует реабсорбции воды в дистальных отделах нефрона путем увеличения проницаемости для воды стенок дистальных извитых канальцев и собирательных трубочек. Мех-зм действия АДГ заключается в активации фермента аденилатциклазы, который участвует в образовании цАМФ из АТФ. При избытке АДГ может наступить полное прекращение мочеобразования. Уменьшение секреции АДГ вызывает развитие тяж. заболевания несахарного диабета. При этом заболевании выделяется большое кол-во светлой мочи с незначительной относительной плотностью. АДГ имеет важное значение в поддержании осмотического давления крови. Альдостерон увелич-т реабсорбцию ионов натрия и секрецию ионов калия и водорода клетками почечных канальцев. Одновременно возрастает реабсорбция воды, которая всасывается пассивно по осмотическому градиенту, что приводит к уменьшению диуреза. Гормон уменьшает реабсорбцию кальция и магния в проксимальных отделах канальцев. Натрийуретический гормон усиливает выведение ионов натрия с мочой. Паратгормон стимулирует реабсорбцию кальция и тормозит реабсорбцию фосфатов. Кальцитонинтормозит реабсорбцию кальция и фосфата. Адреналин суживает просвет выносящих артериол, в результате чего повышается гидростатическое давление, увеличиваются фильтрация и диурез. Инсулин. Недостаток этого гормона приводит к гипергликемии, глюкозурии, увеличению осмотического давления мочи и увеличению диуреза. Тироксин усиливает обменные процессы, в рез-те возрастает кол-во осмотически активных веществ, что приводит к увеличению диуреза. Простагландиныугнетают реабсорбцию натрия, стимулируют кровоток в мозговом веществе почки, увеличивают диурез.
96. Регуляция обмена углеводов в организме.Все эти обмены связаны м/д собой ч/з промежуточные продукты. Так, при расщпл-ии гл-зы образ ацетил-коА который далее может использ-ся для синтеза жирных к-т и липидов, а также аминок-т и белков. Т.о. стимул распада углеводов вызывает активацию синтеза белков и жиров. Активация гликолиза или распада аминок-т может стимулир-ть глюконеогенез. Следоват-но обмен углеводов ,белков и жиров регул-т один и тот же набор гормонов. 1)Соматотропный гормон (анаболический, усиливает приток в клетку амикнок-т, и увел. синтез белка). Он действ-т липолетически. Высвобождающиеся жирн.к-ты использ-ся как источник углерода и Е для синтеза белков, увелич. чувств-тьадипоцитов к липолетическому действию катехоламинов. Действие СПГ на углев. обмен, явл бимодальным: может действ-ть на Б-клетки подж/ж-зы и стимул-т секрецию инсулина. Т.о. в 1 фазу наблюд-ся инсулиноподобное действие на углеводн. обмен, во 2 фазу гормон явл контринсулярным; распад и захват гл-зы тормозится, запуск-ся глюконеогенез стимулир-ся гликолиз(диабетическое действие). 2)Глюкокартикоиды (кортизол, кортикостероиды). Кортизол стимулир. в печени глюконогенез, тормозит транспорт гл-зы и ее утилизацию, повыш-т ее концентр-ю в крови. Глюкокортикоиды стимул-т расщепления белков, катаболически действ-т на м-цы, лимф ткань, кожу и кости. У картизола липолетическое действие, что повыш-т концентр-ю свобод-х жир к-т, тормозится утилизация гл-зы и липогенез. Секреция кортикостероидов регул-ся гипофизарным гормоном АКТГ. Глюкортикоиды снижают имунный статус, ослабляют клеточн. иммунитет, блокир-ся высвобождение цитокина и тормозят все воспал. процессы. 3)Гормоны щитов/ж-зы. Усилив общий обмен в-в, увел потребл-е кислорода, Т3 –стимул-т все этапы обмена углеводов: всасыв-е в ЖКТ гликогенолиз, глюконеогенез в печени, жир-й тк. и в мышцах , поддерж-т гликолиз, способствуют липогенезу печени. 4)Инсулин пониж концентрацию гл-зы в крови и явл запасающ гормоном в орг-ме. Инсулин ингибирует гормон чувствит-ю липазу, высвобождающиеся жир к-ты накаплив-ся в печени в составе триглицерина ,что и объясн-т ожирение печени при сахарн. диабте, сниж-ся бета-окисление, повыш-ся синтез жиров. 5)амилин- пептидный гормон (37 аминок-т), синтез-ся в Б-клетках с инсулином и имеет сходный мех-зм секреции (стимулир-ся глюкозой), замедл-сяпоступл-е гл-зы в кровь после приема пищи и подавляет секрецию глюкагона. 6)глюкагон-явл.прямым антагонистом инсулина в печени и частично в жировой ткани. Синтез-ся в А-клетках островков Лангерганса. Выдел-ся при поддержании концентрации глюкозы в крови, обеспечив. снабжение ткани, особенно мозга глюкозой и кетоновыми телами. 7)Катехоламины - индуцируют расщепл-е гликогена в печени и мышцах. Способствуют глюконеогенезу в печени ,стимулир-т липолиз в жировой ткани и протеолиз в печени. Угнетают секрецию инсулина, тормозят все инсулин зависимые процессы, повыш-ся уровень гл-зы в крови. Гипергликемия - симптом, обозначающий увеличение содержания глюкозы в сыворотке крови по сравнению с нормой в 3,3—5,5 ммоль/л. Гипогликемия - состояние, при котором содержание сахара (глюкозы) в крови ниже нормы.
97. Гормональная регуляция обмена углеводов, белков и жиров. Все эти обмены связаны м/д собой ч/з промежуточные продукты. Так, при расщпл-ии гл-зы образ ацетил-коА который далее может использ-ся для синтеза жирных к-т и липидов, а также аминок-т и белков. Т.о. стимул распада углеводов вызывает активацию синтеза белков и жиров. Активация гликолиза или распада аминок-т может стимулир-ть глюконеогенез. Следоват-но обмен углеводов ,белков и жиров регул-т один и тот же набор гормонов. 1)Соматотропный гормон (анаболический,усиливает приток в клетку амикнок-т, и увел. синтез белка). Его недостаток во взрослом состоянии влияет на развитие атеросклероза ,на мышцы, распределение жира, жировой обмен. Он действ-т липолетически. Высвобождающиеся жирн.к-ты использ-ся как источник углерода и Е для синтеза белков, н увелич. Чувств-ть адипоцитов к липолетическому действию катехоламинов. Действие СПГ на углев. обмен, явл бимодальным: может действ-ть на Б-клетки подж/ж-зы и стимул-т секрецию инсулина. Т.о. в 1 фазу наблюд-ся инсулиноподобное действие на углеводн.обмен, во 2 фазу гормон явл контринсулярным; распад и захват гл-зы тормозится, запуск-ся глюконеогенез. Стимулир-ся гликолиз(диабетическое действие). 2)Глюкокартикоиды (кортизол, кортикостероиды). Кортизол составл 95% глюкокортикоидн актив-ти. Они стимулир. в печени глюконогенез, тормозят транспорт гл-зы и ее утилизацию, повыш-т ее концентр-ю в крови. Глюкокортикоиды стимул-т расщепления белков, катаболически действ-т на м-цы, лимф ткань, кожу и кости. Образующиеся аминок-ты вовлекаются в глюконеогенез печени. У картизола липолетическое действие, что повыш-т концентр-ю свобод-х жир к-т, тормозится утилизация гл-зы и липогенез. Секреция кортикостероидов регул-ся гипофизарным гормоном АКТГ. Глюкортикоиды снижают имунный статус, ослабляют клеточн. Иммунитет, блокир-ся высвобождение цитокина и тормозят все воспал. процессы. 3)Гормоны щитов/ж-зы. Усилив общий обмен в-в, увел потребл-е кислорода, Т3 –стимул-т все этапы обмена углеводов: всасыв-е в ЖКТ гликогенолиз, глюконеогенез в печени, жир-й тк. и в мышцах , дополн-но поддерж-т гликолиз, способствуют липогенезу печени). 4)Инсулин пониж концентрацию гл-зы в крови и явл запасающ гормоном в орг-ме. В печени из главных эффектов явл.активации глюкокиназы, актив-ся фосфофруктокиназа и гликогенсинтетаза. Ингибир-ся ферменты расщепления гликогена (фосфорилазы), в мышцах инсулин активирует транспорт гл-зы. Нервн. клетки не зависят от инсулина, что и объясняет гликогликемический шок (0,5-0,2 г/л при норме 0,8-1 г/л) . Инсулин ингибирует гормон чувствит-ю липазу, высвобождающиеся жир к-ты накаплив-ся в печени в составе триглицерина ,что и объясн-т ожирение печени при сахарн. диабте, сниж-ся бета-окисление, повыш-ся синтез жиров. Побочный эффект – стимуляция транспорта аминок-т. Отдаленый эффект – повышение уровня транскрипции ферментов, катализирующих распад гл-зы. 5)амилин- пептидный гормон (37 аминок-т), синтез-ся в Б-клетках с инсулином и имеет сходный мех-зм секреции (стимулир-ся глюкозой), замедл-ся поступл-е гл-зы в кровь после приема пищи и подавляет секрецию глюкагона. 6)глюкагон-явл.прямым антагонистом инсулина в печени и частично в жировой ткани. Синтез-ся в А-клетках островков Лангерганса. Выдел-ся при поддержании концентрации глюкозы в крови, обеспечив. снабжение ткани, особенно мозга глюкозой и кетоновыми телами. Мех-зм действия аденилатцеклазой системы явл кетогенным и гипергликемическим гормоном, активирует гликогенфосфорилазу, ингибирует глюкогенсинтетазу, активирует глюконеогенез, стимулир-т бета-окисл-е в печени и жировой ткани. 7)Катехоламины - индуцируют расщепл-е гликогена в печени и мышцах. Способствуют глюконеогенезу в печени ,стимулир-т липолиз в жировой ткани и протеолиз в печени. Угнетают секрецию инсулина, тормозят все инсулин зависимые процессы, повыш-ся уровень гл-зы в крови.
98.Гормоны щитовидной ж-зы и паражитовидных желез.Синтезир-ся главные клетки из аминок-т тирозина, поступают строем по многичеслен.капиллярам, образующим сеть вокруг скопления тиреоцитов. В тироцитах происходит синтез тироглобулина-белка предшеств. Тироциты поглощают йод. Тиреоглобулин перемещается в полость фолликула. Там происходит йодирование остатков тирозина и образование монойодтиронина и дийодтиронина. Затем происходит перенос одного йодированного остатка на другой. В таком виде белок образует коллоид и накапливается в фолликуле. По мере необходимости из коллоида захватывается часть белкового комплекса и путем эндоцитоза переносится в лизосомы, в котор. происходит отщепление йодированных остатков и образование активных форм гормона. Активная форма транспортируется в капилляры, секретируется в кровоток и присоединяется к транспортному тиронин-связывающему белку.Гормоны щит. ж-зы необходимы для норм.развития человека. Действие гормонов щ/ ж-зы. В перината/льном периоде-формируется НС и скелет. В перинатальном и постнатальном периоде: 1.Увелич-е потребления О2 в тканях, кроме мозга, селезенки и яичек. 2.Регул-я деятельности дых.центра. 3.Стимул-я эритропоэза. 4. Повыш-е теплопродукции. 5. Положит-е хронотропное действие на миокард(отриц. действует на фермент АТФазу, обычно перекачивающий Na из клетки,а H-в клетку. Вследствие обратного тока иона происходит накопл-е Ca в клетке, активация сокращения, сужение комплекса УРСТ на ЭКГ. 6. Повыш-е чувствительности к котехоламинам. 7. Повыш-е кол-ва котехолов. Рецепторов в организме. 8. Повыш-е оборота костной ткани(стимуляция и реабсорбция и образование кости). 9. Стимулирует функцию желтого тела(поддержание беременности на ранних сроках). Синтез гормонов щ/ж-зы регулируется при помощи гипоталамо-гипофизар. системы. В гипоталамусе синтез-ся тиреоглобулин,перемещаемый при помощи гипоталам-аденогипофизарной трансп-та в переднюю долю гипофиза,где активируется синтез тиреотропонина. Он с кровью перемещается к тироцитам и активирует синтез Т3 и Т4. Секреция териолибирина регул-ся по мех-зму отрицательнообратной связи. Гипотиреоз - заболевание, возникающее, в результате недостаточного обеспечения органов и тканей гормонами щитовидной железы из-за снижения уровня продукции тиреоидных гормонов. Различают первичный (при поражении щитовидной железы) и вторичный (при поражении гипофиза и/или гипоталамуса) гипотиреоз. Симптомы: снижение АД, отечность кожи, снижение аппетита, бледное лицо, нарушение нервн.системы. Гипертиреоз или тиреотоксикоз – это синдром, вызванный гиперфункцией щитовидной железы, т.е. избыточной выработкой тиреоидных гормонов. Симптомы: истонченная, потливая кожа; одышка, выпячивание глазн.яблока, расстройства ЖКТ, учащенное мочеиспускание. Гормоны паращитовидных желез:паратгормон и кальцитонин. Кальцитонин - одноцепочный полипептид, содержащий 32 аминокислотных остатка. Секреция зависит от концентрации кальция в крови. Мишень гормона – костная ткань. Посредник действия – кальцийзависимая АТФаза. Через нее гормон изменяет работу кальциевого насоса. Кальцитонин понижает концентрацию кальция в крови, подавляя его выход из костной ткани. Паратгормон - одноцепочный полипептид, состоящий из 84 аминокислотных остатков. Мишени гормона – почки, костная ткань, ЖКТ. Гормон понижает содержание фосфора и повышает содержание кальция в крови, способствует высвобождению кальция из костной ткани.
99. Половые гормоны: биосинтез, регуляция биосинтеза. Половой цикл.Половые гормоны-Это гормоны гонад. Главные ф-ции гонад – синтез гормонов и обр-е половых кл. Ф-ции пол.гормонов: 1)регул-я гаметогенеза, 2)регул-я развития вторичных половых признаков, 3)половая диф-ка, 4)регул-я анаболич-х процессов. В яичниках обр-ся яйцекл-ки и жен. пол. Гормоны (ЖПГ) – эстрогены и прогестероны(прогестин – гормон желт. тела), а в семенниках – сперматозоиды и муж. пол. гормоны – андрогены. Андрогены синтез-ся клетками Лейдига в интерстиц-й тк. Кровь содержит как муж, так и ЖПГ, но в разных соотн-х у муж и жен. У муж-н с момента рожд-я до 1 года преоблад-т тестостерон, затем их синтез прекр-ся и начин-ся синтез-ся андростерон. В период пол.созр-я способность семенников синтез-ть тестостерон восст-=ся и до конца жизни он явл основным МПГ. Если во время созревания плода половая диф-ка осущ-сь по жен.типу, то с периода пол.созревания возн-т цикл-ть выработки гормнов. Эстрадиол – синтез-ся в созревшем фолликуле яичника, а прогестерон в желт.теле и плаценте. Совместное действие этих гормонов регул-т половой цикл у жен-н. Синтез и секреция пол.гормонов регул-ся при помощи сложной системы отриц.обратн. связи. В гипоталамусе синтез-ся гонадолибирины (люлибирин и фолибирин) котрые влияя на гипофиз активир-т синтез гонадотропных гормонов: 1)фолликулостимулир-й гормон – гликопротеид, состоящий из 2 субъед-ц. Его синтез в аденогипофизе стимул-ся фоллибирином, ингиб-ся эстрагенами (отрицат. обратная связь). Мишени: у жен-н -яичники, где он инициирует развитие фолликулов, клетки внутр-го слоя которых начин-т продуцировать эстрагены. У муж-н – семенники, где он стимул-т развитие эпителия семявыносящих протоков, появл-е большого числа сперматоцитов на всех стадиях развития, включая зрелые (стимул-я сперматогенеза). 2)Лютеинизирующих гормон – гликопротеид. Состоящий из 2 субъед-ц. Синтез актив-ся люлиберином и тормоз-ся по принципу обратной связи прогестерона. Мишени: у жен-н - зрелый фолликул (графов пузырек), ЛГ обеспечив.окончат-е созр-е, овуляцию и обр-е желт тела. У муж-н -клетки Лейдига, где ЛГ стимул-т обр-е тестостерона и семенники, где он стимул-т рост интерстиц-х клеток. Кроме фолликулостимул-го и лютоинизирующего??? гормонов. К гонадотропным гормонам относят пролактин и хорионич-й гонадотропин (ХГЧ). Т.о. синтез тестестерона актив-ся ЛГ, а тормоз-ся прогистероном, который отриц-но действует на синтез люлиберина. Синтиез эстрадиола ктив-ся фолликулостимул-м гормоном, а тормоз-ся эстрагенами (в том числе с самим эстрадиолом по мех-зму отриц.обратн.связи за счет послед-го тормож-я синтеза ФСГ и фолибирина). Синтез прогестерона актив-ся и ФСГ и ЛГ, а тормоз-ся прогестероном по мех-зму отриц. обратн. связи, за счет торможения синтеза ЛГ в гипофизе и люлиберина в гипоталамусе. Регул-я полового цикла. У жен-н уровень секреции образ-ся в яичниках стероидов меняется в различных фазах менстр-го цикла и зависит от скорости их обр-я в яичниках. Менстр-й цикл (МЦ) обусл-ся сложным взаимодействием гипоталамуса, гипофиза и яичниками. В норме продолж-ть цикла от 30-35 дней ( в ср 28 дней). Его подразд-т на: 1)фолликулярную фазу, 2)лютеинизирующая фаза, 3)менструация. 1 фаза- происх-т рост фолликула, стимулируя ФСГ и ЛГ. Рост осущ-ся под влиянием ФСГ, фолликул синтез-т эстрадиол, который в свою очередь по мех-зму полож. обратн. связи стимулир-т синтез и массированную секрецию ЛГ и переходит к след-й фазе. Содерж-е прогестерона очень мало. 2 фаза после овуляции клетки- гранулезы лопнувшего фолликула лютеиниз-ся и обр-т желтое тело, которое синтез-т прогестерон - основной гормон этой фазы и эстрадиол. Прогестерон необх-м для формир-я эндометрия, обесп-х нужное условие для развития имплантирующего бластоциста. До имплантации интенс-ть синтеза прогестерона желт тела обеспеч-ся за счет действия ЛГ, а если имплантация произошла, то дальнейший стимул прогестерона стимул-ся гормоном плаценты ХГЧ. ХГЧ поддерж-т синтез прогестерона желт тела, до тех пор, пока плацента сама не начнет синтез-ть большое кол-во прогестеронов. При отсутствии имплантации желт тело деградир-т и наступает 3 фаза – менструация. После отторж-я эндометрия наступает нов. цикл, разруш-е пол. гормонов происх-т в печени (обр-ся гидроксистероиды), продукты распада удал-ся с мочой, тестостерон превр-ся в андростерон, эстрадиол в эстрол, прогестерон в прегнандиол.
Дата добавления: 2015-03-11; просмотров: 1084;