Шлеммов канал и коллекторные каналы.

|
| Рис. 3.3.10. Сканограмма вскрытого венозного синуса склеры (шлеммов канал). Стрелками указаны трабекулы |
Венозный синус склеры (шлеммов канал; sinus venosus sclerae Schlemm). Шлеммов канал (Schlemm, 1830) представляет собой узкую трубку или систему трубок длиной 36 мм (рис. 3.3.7, 3.3.10, 3.3.11). Внутренняя ее стенка
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

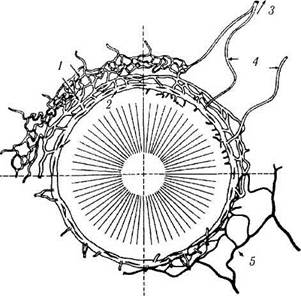
Рис. 3.3.11. Схематическое изображение венозного синуса склеры (шлеммового канала) и его отношение к артериальной и венозной системам (по Tripathi et ai, 1982):
1 — интрасклеральное венозное и глубокое склеральное сплетения; 2 — шлеммов канал; 3 — эписклеральное венозное сплетение; 4 — водяные вены; 5 — артериальный круг
выстлана эндотелием. Шлеммов канал располагается в наружной части внутренней склеральной борозды. Его основной функцией является отведение камерной влаги из трабекулярной сети в эписклеральную венозную сеть посредством коллекторных каналов. Юкстаканалику-лярная соединительная ткань отделяет внутренние и внешние стенки шлеммова канала от трабекулярной сети и склеры.
Просвет шлеммова канала на поперечном разрезе овальной формы [81]. Он может быть разделен перегородками на отделы и состоять из многочисленных каналов.
Ширина шлеммова канала в поперечных плоскостях 120—400 мкм и 10—25 мкм [253, 1103]. Существуют довольно широкие колебания размеров шлеммова канала в зависимости от возраста, наличия предшествоваших заболеваний глаза, что необходимо учитывать при проведении антиглаукоматозных операций [161, 396, 775, 776, 902].
Эндотелиальная выстилка шлеммова канала располагается на базальной мембране, которая местами прерывается. Подобный характер базальной мембраны позволяет предположить, что мембрана не может обеспечить существенного сопротивления потоку камерной влаги.
Главными компонентами базальной мембраны являются коллаген IV типа, ламинин, фиб-ронектин, гепаран сульфат протеогликан [387, 389, 697, 766, 1070].
В шлеммовом канале видны отростчатые расширения в виде дивертикулов, направлен-
ные в сторону юкстаканаликулярной ткани и трабекулярного аппарата (каналы Сондерман-на; [1022]).
Эндотелиальная выстилка. Стенка шлеммова канала, обращенная в сторону глаза. На протяжении длительного времени продолжались споры относительного того, существует или нет прямое сообщение между передней камерой и шлеммовым каналам [645, 646, 980]. Теперь точно известно, что передача влаги осуществляется посредством переноса ее через цитоплазму эндотелиальных клеток. Морфологическим проявлением этого процесса является присутствие в цитоплазме эндотелиоцитов вакуолей [154, 1094].
Шлеммов канал выстлан одним слоем эндотелиальных клеток. На внутренней поверхности канала они имеют длину 40—120 мкм, ширину 4—12 мкм, а толщину 0,2 мкм [1103]. Скреплены они между собой при помощи дес-мосом. Встречаются и редкие щелевые контакты, располагающиеся между эндотелиаль-ными клетками и клетками юкстаканаликулярной ткани.
Межклеточные контакты занимают незначительную площадь мембраны. Они не могут предотвратить прохождение лейкоцитов или макрофагов. Плотность расположения межклеточных контактов не изменяется при изменении внутриглазного давления [1207].
На апикальной поверхности эндотелиальных клеток видны микроворсинки. В цитоплазме эпителиоцитов содержатся многочисленные свободные рибосомы и микрофиламенты, а также множество пиноцитозных пузырьков.
Наиболее явной особенностью внутренней стенки шлеммова канала является наличие гигантских вакуолей. Ширина их от 4 до 6 мкм, а длина до 25 мкм. Возникают они в результате инвагинации базальной плазматической мембраны эндотелиальных клеток, обеспечивая, таким образом, возможность проникновения камерной влаги в юкстаканаликулярную ткань [154, 368, 369, 566, 1024, 1093—1099, 1103, 1132, 1133].
Меньшая часть влаги может проникать через поры, образованные в цитоплазме клеток («трансцеллюлярные каналы») [1103]. Поры могут быть до 2,5 мкм в диаметре, в то время как базальные инвагинации имеют ширину до 4 мкм. Плотность расположения пор в норме равняется 850 пор/мм2 (Johnson et al., 2002), причем их плотность уменьшается при развитии глаукомы.
Использование меченных изотопами веществ и частиц различного диаметра позволило выяснить, что многие вещества могут проходить через «трансцеллюлярные каналы» из передней камеры в шлеммов канал (рис. 3.3.12). Эта возможность выявлена для торотраста, ферритина, золота и пероксидазы хрена. Через эти каналы могут проходить даже
Передняя камера и дренажная система


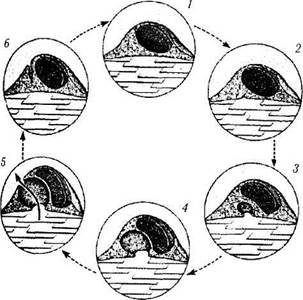
Рис. 3.3.12. Схематическое изображение концепции Tripathi et al. (1977) относительно механизма формирования трансцеллюлярных каналов в эндотелиальных клетках шлеммова канала при выведении камерной влаги (цикл образования вакуолей в эндотелиальных клетках):
] — влага в просвете канала; \-гЩ — влага в межтрабекуляр-ном пространстве
такие клетки, как эритроциты [173, 312, 380, 405—409, 653].
Выявлена закономерность, которая сводится к тому, что формирование вакуолей в эндотелиальных клетках зависит от уровня внутриглазного давления. Причем при нарастании давления число вакуолей увеличивается [405—409, 546, 566, 995, 1095].
Tripathi [1098, 1099] считает, что при увеличении внутриглазного давления в эндотелиальных клетках внутренней стенки шлеммова канала появляется способность «циклически» пропускать камерную влагу, образуя внутрицито-плазматические вакуоли и «трансцеллюлярные каналы» [115, 1098]. До сих пор непонятно, является ли этот процесс активным, использующим энергию, или протекает пассивно. Тем не менее важно знать, что камерная влага поступает в шлеммов канал только через эндо-телиальные клетки и только 1% общего объема влаги проникает между эндотелиальными клетками [407, 408, 877]. При этом вся эндотелиаль-ная выстилка шлеммова канала обеспечивает только 5—10% сопротивления оттоку камерной влаги [115, 295, 406].
Эндотелиальные клетки наружной стенки шлеммова канала более длинные и более плоские. Апикальная их поверхность гладкая. Они прочно соединены между собой при помощи зон замыкания. В цитоплазме клеток редко выявляются гигантские вакуоли. Лежат эндо-
телиоциты на толстой базальной мембране. В соответствии с особенностями строения наружной стенки можно предположить, что ее пропускная способность низкая. Тем не менее использование изотопных меток выявило высокую пропускную способность [1103].
Коллекторные каналы. Коллекторные каналы в количестве 25—35 начинаются у внешней стенки шлеммова канала (рис. 3.3.11). Посредством этих каналов влага оттекает в три венозных сплетения: глубокое, среднее склеральное и эписклеральное. До 8 каналов отводят влагу непосредственно в эписклеральное венозное сплетение. Известны эти каналы как «водяные вены». Они были обнаружены Аше-ром (Ascher) в 1942 году, а их связь со шлем-мовым каналом выявлена Эштоном [80].
При помощи щелевой лампы «водяные вены» видны в виде прозрачных сосудов, содержащих как камерную влагу, так и кровь [384]. Наиболее часто их можно обнаружить субконъ-юнктивально на расстоянии 2 мм от лимба книзу и назально. Перед впадением в эписклераль-ные вены они распространяются на протяжении 1,0—10,0 мм. Коллекторные каналы выстланы эндотелием. Клапаны в них отсутствуют.
Глубоко расположенное склеральное венозное сплетение представлено ветвями передних ресничных вен, которые соединяются со средним склеральным сплетением. При этом в лим-бальной области образуется интрасклеральная венозная сеть. Эта система получает кровь также и от ресничного венозного сплетения.
Из интрасклерального сплетения влага оттекает в эписклеральное сплетение и далее к передним ресничным венам. Эписклеральное венозное сплетение, кроме того, получает кровь от вен конъюнктивы, дренирующих перилим-бальную область.
Кровоснабжение дренажной системы.Кро-воснабжается шлеммов канал сосудами малого круга кровообращения радужки, получающего, в свою очередь, ветви из поверхностных и глубоких ответвлений передних ресничных артерий [344]. Иногда артериолы проходят вблизи шлеммова канала, отделенные от него только адвентицией [85].
Иннервация дренажной системы.Иннервация дренажной системы осуществляется волокнами надресничного и ресничного сплетений, расположенных в области склеральной шпоры.
В трабекулярной сети обнаруживаются как миелинизированные, так и немиелинизирован-ные нервные волокна. Миелинизированные волокна образуют дугу, прилегающую к задней поверхности трабекулярного аппарата. Нервные окончания обильны, как в юкстаканали-кулярной ткани, так и в трабекулярной сети (рис. 3.3.13) [154, 190, 496, 619, 793, 934, 947, 994, 1122, 1137, 1140].
Ruskell [946] нашел немиелинизированные волокна на всем протяжении трабекулярной
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

|
Рис. 3.3.13. Распределение нервных окончаний (треугольники) в области трабекулярной сети и венозного склерального синуса (по Tripathi et al., 1982)
сети и шлеммова канала. Наиболее часто они встречались в юкстаканаликулярной ткани, а также вблизи эндотелиальной выстилки шлеммова канала.
В настоящее время не совсем ясно, к какому типу относятся обнаруживаемые в этой области нервные волокна. Это во многом связано с тем, что нейротрансмиттерами являются многочисленные вещества. Нервные волокна могут быть аминэргическими, нитрэргическими и пептидэргическими [994]. Nomura, Smelser [793], Ruskell [946] считают, что симпатические волокна составляют 30%. Располагаются они в трабекулярной сети и передней части продольной ресничной мышцы и относятся к адренэрги-ческим. Количество подобных волокон уменьшается с возрастом, а также при хронической простой глаукоме [281, 619, 1184]. Некоторые симпатические нервы иммунореактивны к ней-ропептиду Y [139, 1041, 1042].
Парасимпатическая иннервация угла передней камеры посредством волокон ресничного ганглия выявлена Holland, von Salirnan, Collins [498]. Ruskell [946, 935] установил, что у обезьян парасимпатические волокна поступают с лицевым нервом, образующих синапсы в крыло-небном ганглии. В глазницу они поступают посредством rami orbitales [1040].
Имеются данные, свидетельствующие о том, что нервы, исходящие из крылонебного ганглия, иммуноактивны в отношении вазоинтер-стициального полипептида (VIP). Эти волокна также иннервируют заднюю часть увеального тракта глаза человека [1040].
Чувствительные волокна тройничного нерва, содержащие Р вещество, выявлены в структурах угла глаза обезьян и человека, а также в увеальных и корнеосклеральных частях трабекулярной сети, юкстаканаликулярной ткани и шлеммовом канале [1040]. Иннервиру-
ются пептид-, нитр- и аминэргическими волокнами и миоэпителиальные клетки [1062—1064, 1066].
Отдельно необходимо остановиться на меха-норецепторах, обнаруживаемых в дренажной системе. Формируются они следующим образом. Внутренние слои глаза млекопитающих иннервируются сенсорными нервами, исходящими из тройничного нерва. Большинство волокон относится к волокнам типа С [101, 498], а некоторые из них специфически окрашиваются на субстанцию Р [538].
Многочисленные ветви тройничного нерва проникают в склеру. При этом часть миели-низированных волокон образуют склеральное сплетение. Именно от этого сплетения отходят ветви к трабекулярной сети, теряя при этом миелиновую оболочку. Заканчиваются эти волокна нервными окончаниями типа механоре-цепторов [1062]. Рядом исследователей показано, что по строению механорецепторы трабекулярной сети наиболее близки к бароре-цепторам [182, 618, 994]. Эти рецепторы специфически окрашиваются на наличие белков нейрофиламентов и синаптофизин, т. е. маркер синаптических пузырьков [244]. Рецепторы трабекулярной сети подобны висцеральным механорецепторам других частей тела — каро-тидного синуса, дуги аорты, эндокарда, системы органов дыхания, пищевода, кожи, сухожилий [74, 433, 434, 582, 583, 610, 777, 883, 1086].
Количество и плотность расположения меха-норецепторов трабекулярной сети увеличиваются с возрастом [1062, 1123, 1138], а также при хронической простой глаукоме.
Существует три гипотезы, объясняющие роль механорецепторов, расположенных в области дренажной системы [1062]. Они могут выполнять функцию проприоцепции сухожилий ресничной мышцы, влиять на сокращение мио-фибробластоподобных клеток склеральной шпоры [1066, 1068]. Кроме того, они могут функционировать как барорецепторы при изменении внутриглазного давления.
Дата добавления: 2014-12-20; просмотров: 3417;