Эстуарии и затопляемые земли
Научная деятельность океанологов в эстуариях и на затопляемых низменностях еще более специфична, чем прибрежная океанология в целом. Для этого есть несколько причин.
1. В эстуариях соленость быстро изменяется на коротких расстояниях, причем как в горизонтальном, так и в вертикальном направлении. Это означает, что животные должны уметь свободно перемещаться с тем, чтобы сохранить свое положение в области нужной им солености волы и приспособиться к изменению осмотического давления.
2. Характер циркуляции воды в эстуариях заметно изменяется в зависимости от величины речного стока. Для тех мест, где в эстуарии происходит смешение соленых и пресных вод, характерны сильные сезонные изменения.
3. Практически на всех эстуариях сегодня серьезно сказывается влияние человеческой деятельности. Почти невозможно узнать истинные, первоначальные особенности какой бы то ни было экосистемы эстуариев.
4. Многие морские организмы, живущие во взрослом состоянии преимущественно в морской воде, мигрируют в эстуарий и обратно для питания или нереста; так делают северный менхеден и тихоокеанская сельдь. Эпизоды миграции связаны со сменой времен года и начинаются после характерного сезонного изменения какого-то фактора, например притока пресной воды, температуры или солености (рис. 12.7).
Циркуляция воды в эстуариях. Процессом перемешивания пресной и морской воды в эстуариях управляют два фактора: величина океанского прилива в устье эстуария и величина стока рек в вершине эстуария. Перемешивание в эстуарии может быть двух типов: полным, когда определяющую роль играет величина прилива, или в виде клина соленых вод, когда доминирует речной сток.
«Полностью перемешанный» эстуарий. Если велики океанские приливы, или слаб приток речных вод, или же если удовлетворяются оба этих условия, то вода в эстуарии перемешивается довольно хорошо сверху донизу, а соленость плавно изменяется от нормальной для океана величины около 30%о в устье эстуария до нуля в его вершине.
Рис. 18.18, а иллюстрирует это положение для эстуария реки Колумбия в такое время года, когда речной сток спадает до минимума, а тихоокеанские приливы в устье эстуария высоки.
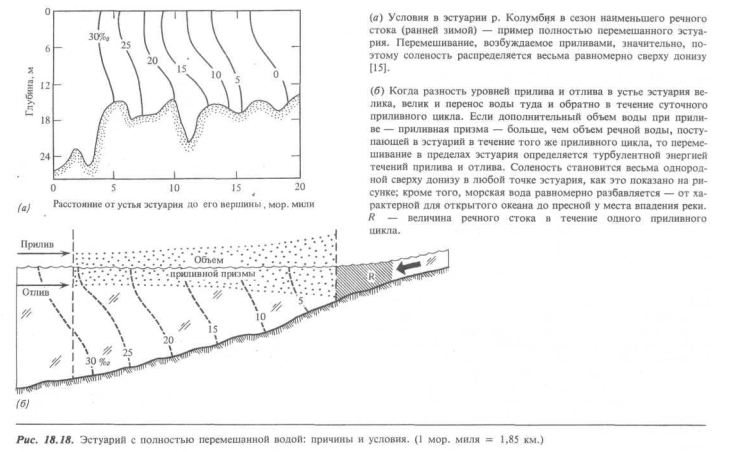
Энергия для вертикального перемешивания обеспечивается турбулентностью, создаваемой внутри эстуария приливными течениями; чем больше скорость приливного течения, тем выше степень турбулентного перемешивания в толще воды.
Можно также рассмотреть этот тип эстуарного перемешивания с точки зрения количества пресной воды, поступающей из реки за время приливного цикла (рис. 18.18, б). Вода, поступающая из прибрежной части океана за этот цикл, должна занимать объем между уровнями прилива и отлива, называемый приливной призмой. В данном случае ясно, что приливная призма существенно больше объема речного стока, поэтому приливные течения бывают сильными. Турбулентность, создаваемая приливными течениями, разрушает естественное плотностное расслоение, которое в противном случае оказалось бы сильнее: пресная вода распространялась бы от реки над слоем более плотной морской воды. В некотором смысле можно сказать, что водный столб в полностью перемешанном эстуарии динамически дестабилизируется снизу.
В редких случаях входящая в эстуарий приливная волна полностью турбулентной мошной стеной мутной воды устремляется вверх по течению, создавая гидравлическую волну, которую называют бором.
Эстуарий с соленым клином. В другом крайнем случае главную роль в процессах перемешивания в эстуарии играют большие массы речной воды. Величина приливов может быть большой или малой, но сама речная вода «одержит основную долю энергии движения в турбулентной форме. Когда пресная вода встречается с соленой водой и перекрывает ее, турбулентные вихри захватывают «порции» соленой воды и переносят их вверх, в лежащий выше слой пресной воды. Этот тип перемешивания, при котором поверхность раздела между двумя различными жидкостями эродируется только с одной стороны, называется вовлечением. Как будет показано в дальнейшем, этот процесс оказывает большое влияние на поведение прибрежных экосистем.
В процессе вовлечения (рис. 18.19) некоторое число R единичных объемов пресной воды попадает в эстуарий в его вершине; значит, такое же число R этих объемов должно выйти из устья эстуария за то же самое время.
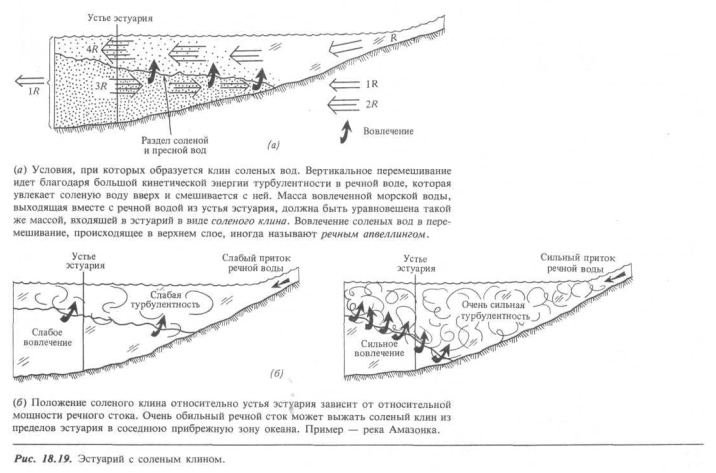
Но из-за того что речная вода осолоняется при своем движении, полный объем солоноватой смеси в устье эстуария должен быть гораздо больше, чем R единиц. Например, если 1 R речной воды вовлекает в процесс перемешивания 3 R единиц морской воды, то для сохранения баланса пресной воды в эстуарии в море должно выйти в общей сложности 4 R единичных объемов солоноватой воды. Но 3 R единиц морской воды в этой солоноватой смеси должны быть скомпенсированы соответствующим притоком из моря. Это объясняет особый характер эстуария с соленым клипом, а именно то, что морская вода втекает внутрь эстуария вдоль дна, восполняя непрерывную потерю морской воды через раздел с турбулентной речной водой. «Клин» морской воды проникает на некоторое расстояние вверх по течению — отсюда и название этого типа эстуариев.
Где в эстуарии располагается соленый клин? Находится ли вершина клина около устья эстуария или проникает далеко внутрь против течения? Ответы зависят от двух факторов — величины речного стока и формы самого эстуария. Сток реки Амазонки столь велик, что клин соленых вод буквально полностью выжат из устья эстуария; океанологами изучен протяженный шельф солоноватых амазонских вод в прибрежной области океана, растекающихся далеко от устья эстуария. Однако у большинства рек вершина клина располагается в пределах эстуария (рис. 18.19, б). В первые месяцы лета, когда сток максимален, клин ближе к устью эстуария, но выражен тоже очень отчетливо; скорость процесса вовлечения и компенсирующий поток морской воды, направленный вверх по эстуарию, велики. Когда величина речного стока уменьшается в осенний сезон, сокращается и энергия турбулентности, питающая процесс вовлечения; клин соленых вод продвигается тогда выше по течению.
Форма эстуария определяет тип циркуляции, влияя на «быстроту» течения речной воды, а значит, и на степень турбулентности в ней. Реки с узкими эстуариями и большим расходом воды вытесняют клин к устью эстуария. Широкие реки, даже с мощным стоком, как правило, менее турбулентны, так что клин соленых вод располагается там ближе к вершине эстуария.
Классический пример эстуария с соленым клином — эстуарий реки Миссисипи. В Мексиканском заливе прилив невысок — 0,5 м или меньше, объем речного стока очень велик, а река весьма широкая. Клин соленых вод распространился вверх по течению до Батон-Ружа, находящегося на расстоянии 200 км от устья реки.
«Частично перемешанный» эстуарий. Характер циркуляции в большинстве эстуариев представляет собой нечто среднее между описанными экстремальными случаями. Турбулентная энергия, питающая процесс перемешивания, распределена в той или иной степени между слоем речной воды и продвигающимся против течения придонным потоком соленых вод. Иначе говоря, соленость верхнего слоя в пределах эстуария заметно растет по мере его движения к устью, а придонный слой непрерывно разбавляется пресной водой по мере своего перемещения в противоположном направлении.
Прекрасный пример частично перемешанного эстуария представляет собой система Чесапикского залива. Эстуарий здесь заходит в пределы континента на 300 км и на этом пути принимает сток многих рек. Воды его относятся к категории солоноватых; изогалина поверхностного слоя 5%о в конце лета проходит очень близко к вершине эстуария (рис. 18.20).
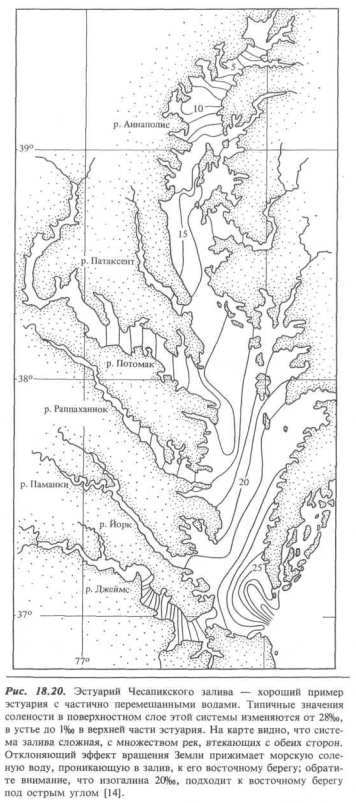
Обратите внимание также на то, что изогалины поверхностных вод ориентированы под косым углом к оси русла, в результате чего создается впечатление, что направленный против течения поток соленой воды распространяется вдоль восточного (правого по отношению к этому потоку) берега, тогда как текущая к морю солоноватая вода прижимается к своему правому берегу. Это действительно так: сила Кориолиса, создаваемая вращением Земли, отклоняет оба потока вправо от направления течения.
Связь прибрежных экосистем с эстуариями. Площадь США включает около 15 150 км2 эстуарных вод. В 1970 г. эти воды вместе с водами шельфа дали 5 млн. т рыбы и других морских продуктов [17]. Мы знаем теперь, что многие виды животных, добываемые на шельфе для продажи, в течение некоторой части своего жизненного цикла не могут обойтись без захода в эстуарии. Мак-Хью [14] подсчитал, что 66% промысловых видов на атлантическом шельфе США зависят от эстуариев, а для отдельных районов процент должен быть еще выше. Используя эти значения, можно вычислить степень такой зависимости: на 1 км2 поверхности эстуарных вод соответствующий региональный промышленный улов составляет 200 т/год. Значит, каждый квадратный километр эстуарных вод, потерянный в результате засыпки грунтом («отвоеванные у моря» площади) или загрязнения, понижает уровень годовой добычи рыбы на 200 т!
Какие же особенности вод эстуариев столь необходимы для прибрежных морских экосистем и как эти особенности связаны с прибрежной фауной? Ответы на оба вопроса связаны с двумя главнейшими отличительными чертами эстуариев: чрезвычайно высокой первичной продуктивностью их вод и своеобразной двухслойной и разнонаправленной системой циркуляции. Более близкий к нашей теме вопрос — почему в эстуариях столь велика первичная продукция? Ответ на этот вопрос также имеет две стороны. Во-первых, масса растворенных питательных веществ и частиц органического материала, заключенная в речной воде, обеспечивает водорослевую продукцию в самой этой речной воде. Во-вторых, вызываемое турбулентностью вовлечение морской воды из клина соленых вод — своего рода апвеллинг, возбуждаемый реками, — «выкачивает» богатую питательными веществами морскую воду с глубины наверх, в эвфотическую зону.
Возбуждаемый реками апвеллинг: пример реки Колумбия. Процесс вовлечения, развивающийся на поверхности клина соленых вод, приводит к тому же биологическому результату, что и апвеллинг, обусловленный ветрами у открытого побережья, а именно увеличению первичной продуктивности. Прекрасный пример этого дает эстуарий реки Колумбия. На рис. 18.21, а показано, как вследствие вовлечения глубинная океанская вода подкачивается в эстуарий.

Сама река приносит в эстуарий мало свободных нитратов, так как они уходят на рост водорослей в реке. В глубинной же океанской воде содержание нитратов, как показано на рис. 18.21,6, очень велико — более 30 мкг-атом/л. Область эстуария, в которую вовлекаются эти нитраты, потенциально весьма продуктивна, что подтверждается схемой, представленной на рис. 18.21, в.
Однако необходимо подчеркнуть одно важное различие между апвеллингом в эстуарии и механизмами прибрежного апвеллинга в океане. В эстуарии — вне зависимости от интенсивности вовлечения морской воды — солоноватая смесь всегда остается менее плотной, чем подстилающая ее океанская вода прибрежной зоны, поэтому находящиеся в эстуарной воде питательные вещества остаются в освещенной солнцем зоне вплоть до истощения. Наоборот, возбуждаемый ветрами апвеллинг выталкивает плотную глубинную воду прямо на поверхность; прекращение ветра позволяет этой воде опуститься обратно и выйти из эвфотической зоны еще до того, как ее питательные вещества будут полностью использованы. Кроме того, ветры имеют переменный характер, а турбулентный речной поток возбуждает процесс вовлечения днем и ночью непрерывно в течение всего сезонного цикла стока.
Продуктивность эстуариев можно оценить, сопоставив максимальное ее значение, полученное для реки Колумбия (рис. 18.21, в) и равное 20 мг С/(м3 • ч), с типичным значением для прибрежного апвеллинга — 300 г С/(м2 • год), указанным в табл. 1.2. Чтобы выразить эти значения в сравнимых единицах, предположим, что продуктивный сезон для реки Колумбия длится 6 мес по 10 ч в сутки (т. е. только в дневные часы), и рассмотрим слой толщиной 10 м. В этом случае суммарная продукция составит 900 г С/(м2 • год), или 9 т С/(га • год), т. е. втрое больше, чем при апвеллинге в прибрежной части океана. Интересно сопоставить эти значения с урожаями «сухопутных» злаков. Хорошая ферма Айовы производит в год 3 т обмолоченного зерна с гектара; если кормить им птицу, то это даст около 1 т куриного мяса. В эстуарии же превращение растительной пищи в зоопланктон имеет эффективность 20%, а зоопланктона в промысловую добычу (такую, как улов менхелена) — тоже 20%, что означает превращение 9тС/(га-год) приблизительно в 900 кг рыбы на гектар, т. е. примерно столько же, сколько весят цыплята, вскормленные айовской нивой. Но эстуарии дают не только рыбу: там ведется также промысел креветок и других морских животных, так что общий итог выше, чем для фермы в шт. Айова.
Эстуарии как место нереста и вскармливания молоди. При такой высокой продуктивности эстуарных вод неслучайно, что многие виды фауны прибрежной области океанов приспособились к обитанию в этих богатых водах в течение какой-то части или даже всего жизненного цикла. На рис. 18.22 выделены четыре категории нектона, обнаруженные в эстуарных водах и различающиеся по степени и характеру своей зависимости от этой зоны.

Несколько видов пресноводных рыб было встречено в солоноватых волах; предполагается, что видов, эндемичных для эстуариев, очень мало. Подавляющее большинство видов рыбы, пойманной когда-либо во внутренних водах эстуариев, и определенно все те виды, которые служат объектом промыслового лова, проводят в эстуарии только какую-то часть своего жизненного цикла. Они делятся на два типа: анадромно-катадромные, т. е. виды, проходящие через солоноватые воды эстуариев по пути из пресных в соленые воды или наоборот, и виды, характерные для открытого океана, мигрирующие в эстуарные воды либо в поисках пищи, либо для нереста.
Особый характер циркуляции в эстуариях, при которой морская вода втекает в соленый клин и выходит из него вследствие вовлечения в речную воду, создает механизм, посредством которого личинки и молодь многих видов животных могут кормиться благодаря высокой продуктивности эстуарной зоны в течение гораздо более продолжительных периодов времени, чем обычное время смены вод. Этот механизм иллюстрируется на рис. 18.23 как для нерестящихся, так и для кормящихся в эстуариях видов.
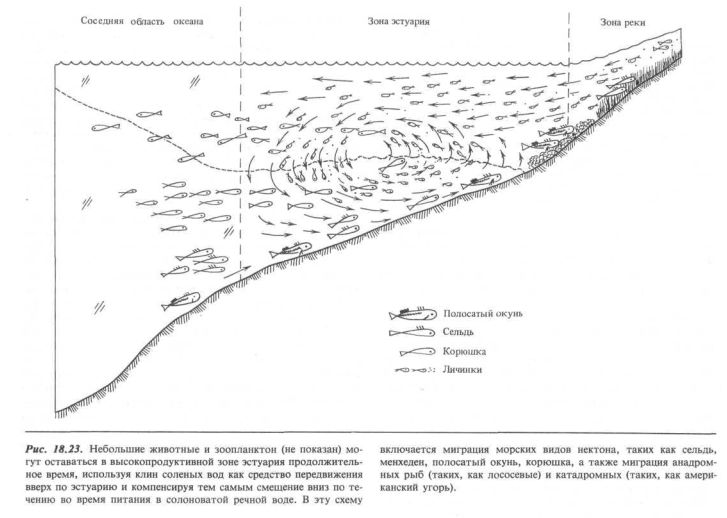
Тихоокеанская сельдь заходит в эстуарии вплоть до литорали, где нерестится, оставляя тяжелые сгустки липкой икряной массы, которая прикрепляется к болотным или водным травам, таким как взморник (Zostera из семейства наядовых). Атлантическая корюшка движется далеко вверх против течения и откладывает икру на дне участков рек с быстрым течением, американская сельдь также мечет икру на речное дно, но в более спокойных водах небольших речек и прудов в верхних частях речного бассейна. Полосатый окунь оставляет икряную массу сразу же над поверхностью распространения соленых вод. Хейдрих и Холл описывают поведение рыб следующим образом [8]:
«Рыба нерестится в конце мая или начале июня над уровнем соленых вод [рис. 18.23]. Полупелагические икринки и личинки проводят, по-видимому, несколько дней или недель почти у самого дна реки, слегка дрейфуя, если течение у дна не слишком сильное. Примерно в двухнедельном возрасте мальки встречаются уже по всей глубине водного столба и начинают дрейфовать по течению. Становясь достаточно сильной для первых попыток свободного плавания, рыба начинает мигрировать по вертикали, перемещаясь ближе ко дну в дневное время и ближе к поверхности ночью. Оказавшись в зоне влияния эстуарного соленого клина, рыбная молодь ночью выносится в сторону моря потоком относительно пресной поверхностной воды, а днем, когда она ближе ко дну, глубинный поток несет ее вверх по эстуарию. Приблизительно в шесть недель рыба уже может сохранять свое положение, плывя против течения. Прямое соперничество (между видами) исключается благодаря разным местам срокам нереста».
Однако независимо от того, производится ли икрометание во внутренних водах или в море, время, когда рыбная молодь в большом количестве достигает эстуария, почти точно совпадает с периодами максимума пищевой продукции, в чем заключается, очевидно, стратегия эволюции.
Сложность экосистемы, включающей условия реки, эстуария и прибрежной зоны океана. Теперь уже читатель может в какой-то мере понять сложность «прибрежной океанологии» и то, почему данная тема рассматривается в этой книге отдельно. Но имеются и особые факторы, которые необходимо здесь упомянуть: эффект климатических изменений, физиологический выбор между «ценой» и «выгодой» для рыб, обитающихся в водах переменной солености, и, наконец, влияние человеческой деятельности.
Если приток в зону эстуария соленой воды с большим содержанием нитратов увеличивается по мере возрастания речного стока (и турбулентности), то многолетние изменения в гидрологическом цикле региона могут дать толчок и к изменениям в фаунистическом сообществе эстуария и прибрежной области океана. На рис. 18.24 показана хорошая корреляция между годовыми уловами палтуса в канадской провинции Квебек и ежегодными мартовскими изменениями расхода пресной воды в устье реки Св. Лаврентия.
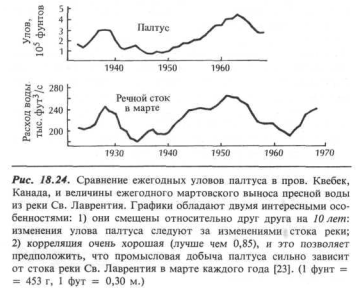
Возрастание мартовского стока отражается в увеличении улова палтуса спустя 10 лет! Практическое использование этих выводов ясно: возможность рациональной организации добычи палтуса на основе прогнозов, сделанных по изучению речного стока.
Если можно сказать, что в результате увеличения расхода пресной воды условия для выживания палтуса становятся более благоприятными, то столь же ясно, что все виды, обитающие в системе эстуария, также испытывают физиологический «стресс», подвергаясь в ходе этого процесса большим изменениям осмотического давления, а кроме того, и тепловому удару, если температура в мелководном эстуарии изменяется быстрее, чем организмы могут к этому приспособиться. Как исследователь оценивает количественные величины, характеризующие эту очевидную связь между ценой усилий и получаемой выгодой? Где находится равновесие между энергетической ценой физиологической адаптации и доступностью пищи при более высокой численности популяции?
Такие вопросы изучены пока недостаточно, но мы должны получить на них ответы. Нужда в ответах на эти вопросы прямо связана с огромным разнообразием условий и органических видов в эстуариях. Множество тонко сбалансированных соотношений разного типа связывает существование специализированных экологических ниш, сроки наступления отдельных стадий гидрологического цикла и другие факторы вроде солености, температуры и мутности воды. Мы уже знаем, что вмешательство людей в жизнь системы река—эстуарий—океан обычно приводит к уменьшению видового разнообразия. Мы обычно изменяем одно или несколько условий и тем самым устраняем те виды, чья жизнедеятельность была в какой-то мере связана с этими условиями. Примеров тому великое множество. Строительство плотин в верхнем течении реки может изменить время возникновения гидрологических импульсов в зоне эстуария (вспомним рис. 12.7). Сброс сточных вод и других отходов воздействует на распределение питательных веществ (например, в Чесапикском эстуарии популяции синего краба заметно выросли в тех зонах, где производится усиленный сброс сточных вод).
Заключение. Что побудило нас включить мини-курс прибрежной океанологии в учебник по общей океанологии? Для этого есть много причин; мы распределим их здесь по отдельным научным дисциплинам.
1. Физика. Трение между материком и движущимися массами океанских вод — главный механизм, компенсирующий тот вращающий момент, посредством которого ветры непрерывно отклоняют течения в океане. Процессом, заставляющим силу трения влиять на характер движения окраинных вод океана, служит турбулентность. Читатель вряд ли сможет вообразить лучшую картину прибрежной турбулентности, чем та, что показана на космическом снимке (рис. 14.8); этот снимок — наглядная демонстрация того, как разные вещества с суши, в том числе вода, попадают в открытое море.
2. Химия. Способность глинистых частиц, находящихся во взвеси в речной воде, переносить на себе различные химические вещества, делает изучение химизма прибрежных вод существенной частью работ по охране окружающей среды. Конечно, химия изучает более широкий круг вопросов, чем только загрязнение среды. Чтобы выяснить пределы развития тех или иных химических процессов в открытом море, мы должны проводить точные измерения потока всевозможных солей с суши в море, а следовательно, измерять скорости осадконакопления, растворения карбонатов на глубине и многое другое.
3. Геология. Выяснить, как пески и другой грубый материал перемешаются вверх и вниз от берега и вдоль берегов, как заносятся в эстуарии и удаляются из них, либо попадая в подводные каньоны, либо задерживаясь на отмелях, как в составе других осадков опускаются на дно окраинных областей океана, — все это важные задачи геологического изучения прибрежной области.
4. Биология. Именно биологические аспекты изучения прибрежной зоны океана особенно сложны. Девяносто девять процентов пищевых продуктов, добываемых в океанах, приходится на их окраины, поэтому необходимо изучить сложные циклы развития всех форм жизни в прибрежной области. Эти циклы тесно связаны с физическими процессами перемешивания на шельфе и в эстуарии. Термические фронты, образующиеся там, где хорошо перемешанный верхний слой воды встречается с перемешиваемым приливами нижним слоем, непосредственно влияют на поведение крупных подразделений в сообществах прибрежной зоны. Биологи пристально изучают и донный субстрат, где на илистом дне, на песке и камнях обитают различные сообщества организмов. Многие аспекты загрязнения, исследуемые химиками, в конце концов оказываются неотделимыми от понимания особенностей существования биологических сообществ.
5. Сравнительно новая отрасль прибрежной океанологии — изучение эстуариев. Зона эстуария весьма важна для прибрежных сообществ, так как она представляет собой место для вскармливания молоди, для питания и размножения различных организмов, а также путь для перемещения многих видов, обитающих в прибрежной зоне, в том числе и для Homo sapiens. Циклические процессы перемешивания, изменения продуктивности и осадконакопления — сложный результат взаимодействия между речными потоками и приливными течениями.
Дата добавления: 2019-10-17; просмотров: 2930;