Соотношение между распределениями по вертикали света и первичной продукции
Зная, что количество световой энергии быстро уменьшается с глубиной, мы можем ожидать, что растения лучше всего развиваются на поверхности моря, где свет наиболее интенсивен. Тем не менее мы обнаруживаем, что это не так. Наблюдениями давно установлено, что больше всего фитопланктона сосредоточено на глубинах примерно от 7 до 17 м. Чем можно объяснить это явное противоречие между потребностями растений и обеспечением их светом?
Объяснение должно включать физический фактор вертикального движения в приповерхностном слое. Вспомните, что конвекция, обусловленная охлаждением и испарением на поверхности океана, вызывает вертикальное перемешивание; дополнительное перемешивание обеспечивают ветровые волны, которые разрушают и создают турбулентное движение в приповерхностных водах. Все движущиеся воды несут с собой мельчайший фитопланктон, так что какое-то вертикальное распределение растительных клеток неизбежно.
С другой стороны, питательные вещества поступают снизу, из-под перемешанного поверхностного слоя. В пределах огромного большинства океанских акваторий, где не происходит апвеллинг, единственный механизм пополнения запасов пищи — это медленная диффузия богатых питательными веществами вод вверх, в поверхностный слой. Турбулентному движению в поверхностном слое способствует вода, активно поступающая снизу. Мы называем этот процесс турбулентной диффузией.
Следовательно, условия, необходимые для возникновения оптимальной продукции растений, должны существовать на каком-то уровне между поверхностью моря, где выше всего интенсивность света, и низом перемешанного слоя, куда поступают питательные вещества. Интервал от 7 до 17 м, в котором мы обнаруживаем самую высокую концентрацию растений, является в среднем компромиссным уровнем, выбранным самими растениями. Вообще при лабораторных экспериментах обнаружилось, что многие растения являются фотоингибиторными, т. е. их продукция снижается, когда они подвергаются воздействию сильного света. Однако замедление развития под действием света свойственно не всем растениям; между видами существует значительный разброс по той интенсивности освещения, при которой каждый вид достигает оптимальной продукции. Например, динофлагелляты, по-видимому, более терпимы к сильной освещенности, чем диатомеи, так что вклад первых в общую продукцию растений гораздо выше в тропических водах. Диатомеи преобладают в более холодных водах высоких широт, где из-за сравнительно меньшей высоты Солнца над горизонтом максимальная интенсивность освещения ниже.
Способность предсказывать первичную продукцию — задача, которую давно пытаются решить биологи-океанологи. Полезным средством для этого является имитационная модель, в которой выделены факторы, предположительно влияющие на поведение растений, а сама модель представляет собой систему математических уравнений, описывающих реакцию растений. Если найдена модель, точно имитирующая реакцию растений в реальном океане, она может работать снова и снова, всякий раз с другим набором весов, приписываемых используемым факторам. Одна такая модель приведена на рис. 14.4.
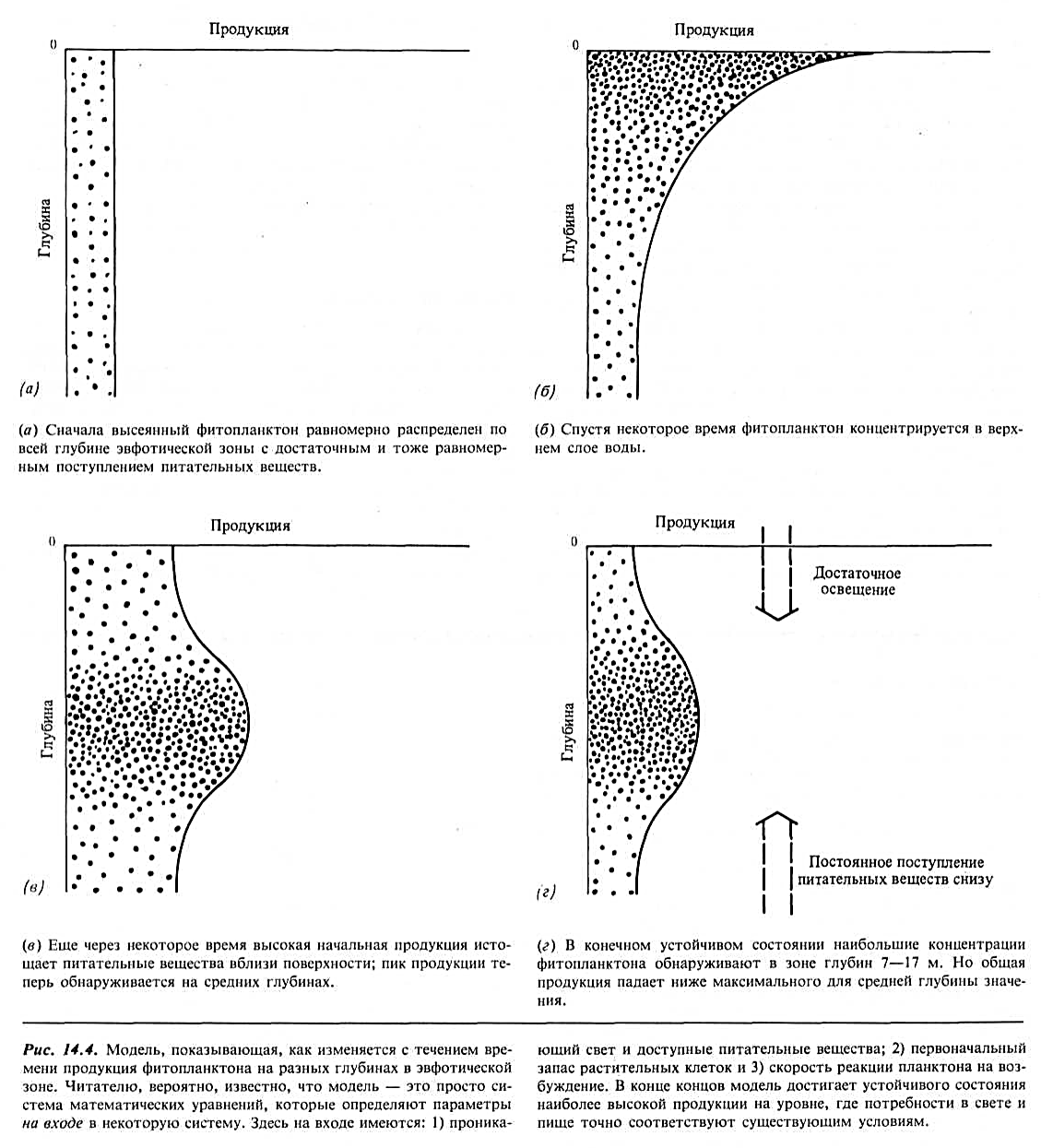
Она начинается с равномерно распределенного планктона, который затем подвергается воздействию изменений в освещенности, обеспечении питательными веществами, выедании и смертности. В результате возникает конечное устойчивое состояние, по которому можно точно предсказать среднее распределение растительных клеток. Такие океанологичекие исследования продолжают проводить, пока еще не разработана универсальная модель.
Глубина перемешивания и точка компенсации.Предполагая, что в целом скорость фотосинтеза падает по мере уменьшения с глубиной интенсивности света, морские биологи разработали понятие точки компенсации. Благодаря тому что скорость дыхания растительной клетки сравнительно постоянна и не зависит от того, продуцирует растение новые клетки или нет, на кривой выделения кислорода должна существовать точка, в которой расход кислорода при дыхании равен образованию его в процессе фотосинтеза. Это точка компенсации, а глубина, на которой она находится, называется глубиной компенсации. На большей глубине в растительных клетках все еще может происходить фотосинтез, но масса клеток все время сокращается, состояние «здоровья» постепенно ухудшается, а вместе с этим снижается и их способность к размножению или к запасанию жира, благодаря которой они сохраняют плавучесть (рис. 14.5, а).
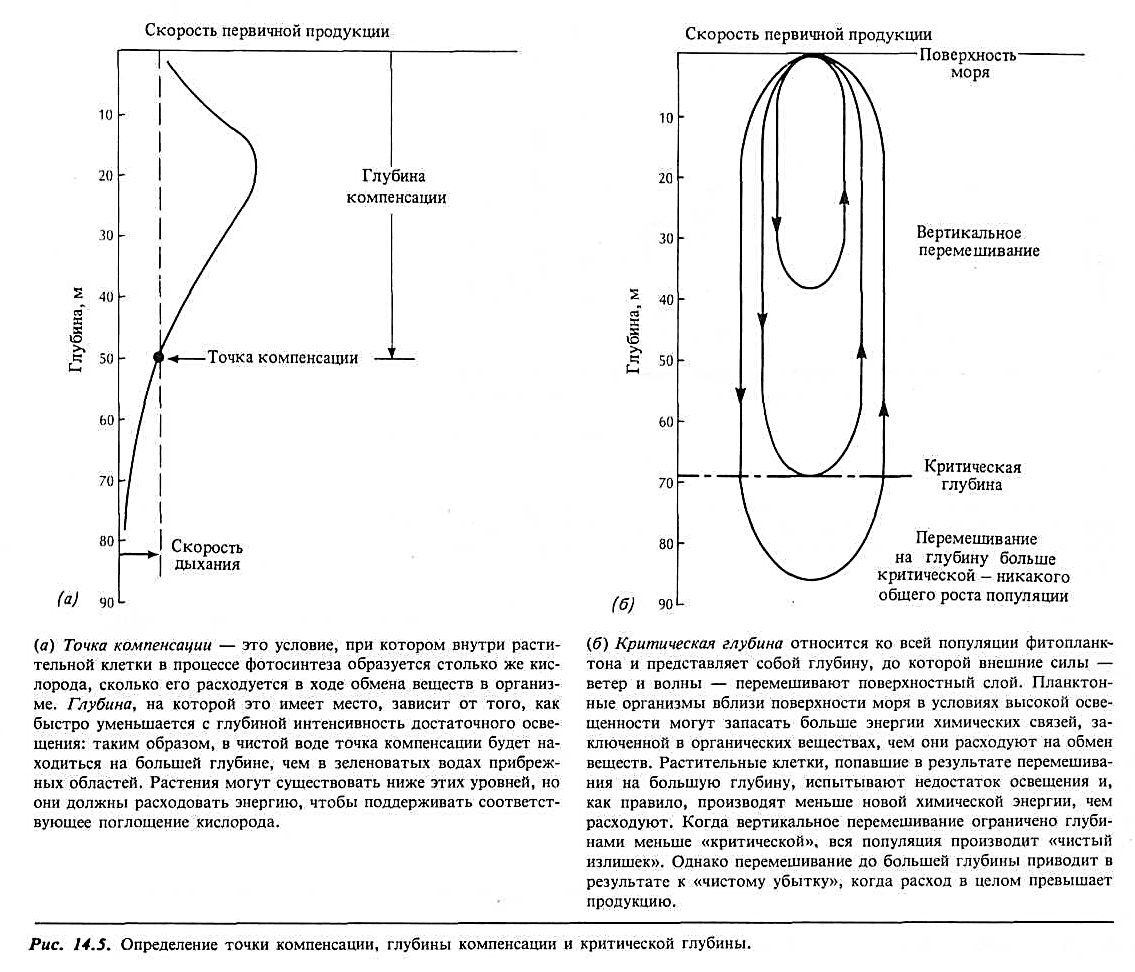
Однако при активном вертикальном перемешивании точка компенсации может приобретать и другой смысл. Если на минутку отбросить все остальные факторы, планктонные организмы должны проводить по крайней мере столько же времени выше этого уровня, сколько и ниже его. Или же они могут приспособиться «использовать» время, которое они проводят выше точки компенсации, для ускоренного синтеза, а затем, пока организм находится ниже точки компенсации, — просто отдыхать или выполнять другие необходимые для клетки функции, например ассимилировать питательные вещества, чтобы использовать их позднее.
Понятие «критической» глубины перемешивания.Глубина компенсации утрачивает свое значение, когда мы имеем дело со всей популяцией фитопланктона. Для этого случая мы вводим другое понятие, которое называется критической глубиной перемешивания. Это глубина, до которой турбулентное перемешивание может распространить всю популяцию, прежде чем «чистая» продукция упадет до нуля. Эта идея (рис. 14.5,б) имеет важные следствия. Если активное перемешивание внутри поверхностного слоя океанских вод происходит до глубин ниже критической, то максимума продукции растений не может быть, так как общая, суммарная потребность в химической энергии для обмена веществ превышает ее производство. Если перемешивание ограничено глубинами меньше критической, имеет место «чистая» продукция, т. е. ее максимум.
Очевидно, что это компромиссная ситуация. Перемешивание поддерживает непрерывное возобновление запасов питательных веществ. Чем больше глубина перемешивания, тем выше скорость, с которой поступают новые питательные вещества, и продукция может возрасти, если достаточно света. Но с увеличением глубины перемешивания планктонные организмы больше времени проводят на глубинах ниже их индивидуальных уровней компенсации и общая продукция может снизиться.
Есть, однако, еще одно условие. Укажем, что чувствительность пигмента, который клетка использует для захвата фотонов, зависит от длины волны. Растение с пигментом, захватывающим фотоны на небольших глубинах, может утратить эффективность поглощения их на более глубоких уровнях из-за того, что длина волны фотона не позволяет ему проникать вглубь, сохраняя достаточную интенсивность. У растений с множеством пигментов, которые покрывают более широкий диапазон поглощаемых длин волн, больше возможностей выжить.
Длина световой волны и чувствительность пигментов.В растительной клетке имеются специальные органоиды, называемые хлоропластами; они содержат хлорофилл и другие пигменты, необходимые для фотосинтеза. Кратко опишем процесс фотосинтеза. Молекула хлорофилла поглощает один или более фотонов определенной длины волны или определенных энергетических уровней и становится «возбужденной» в том смысле, что в пределах этой молекулы один или больше электронов переводятся в более высокое энергетическое состояние. Эти электроны с более высокой энергией проходят в серию других молекул, функцией которых является создание аденозинтрифосфорной кислоты (АТФ); эта кислота в свою очередь служит источником энергии для химической реакции, в результате которой образуются сахара, аминокислоты и другие необходимые молекулы растительной ткани (рис. 14.6, а).
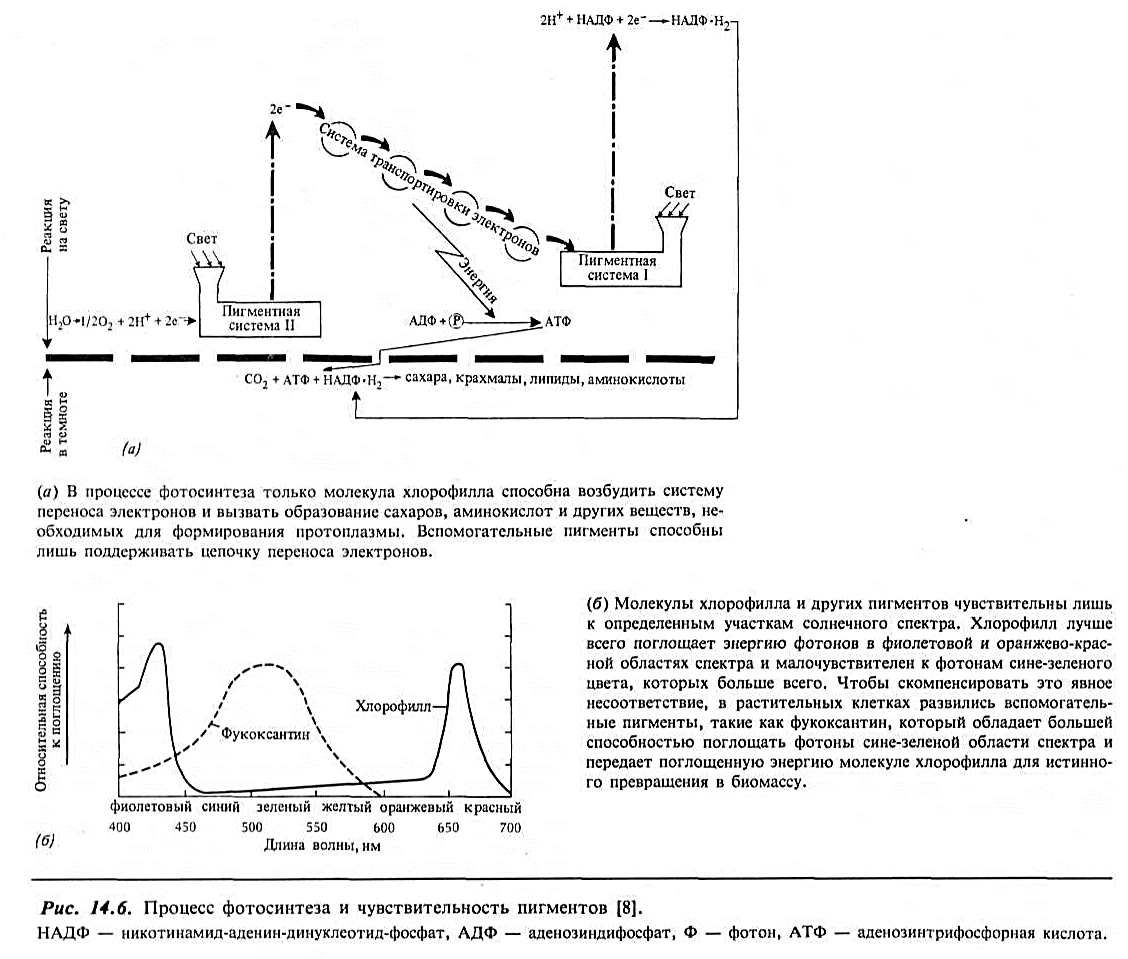
Критическим моментом в этом сложном процессе является «настройка» молекулы хлорофилла на определенные энергетические уровни фотонов. Несоответствие здесь означает, что фотон не поглощается и, следовательно, никакая энергия не производится и не приводит в движение живую систему. Как ни странно, исследования чувствительности хлорофилла к длине волны показывают, что его молекула лучше всего поглощает фотоны, соответствующие фиолетовому (440 нм) и красному (660 нм) цвету (рис. 14.6,6). Из предыдущего обсуждения передачи света в воде повторим, что естественное «окно», пропускающее свет, находится на длине волны около 470 нм в открытом океане и около 500—550 нм в прибрежных водах и в условиях апвеллинга. Фиолетовый и красный свет поглощаются слоем воды толщиной всего в несколько метров. Поэтому, вероятно, может показаться, что растения, зависящие исключительно от хлорофилловых пигментов, окажутся в невыгодных условиях, если попадают на глубину более 5 м.
На рис. 14.6,б показана чувствительность к длине световых волн фукоксантина — вспомогательного пигмента, обнаруженного во многих растениях. Он поглощает энергию в той части спектра падающего света, где сам хлорофилл является плохим абсорбентом. Как и почему возникла такая адаптация? Одна гипотеза основывается на том, что древние океаны представляли собой мелководные моря, где световые волны фиолетовой и красной частей спектра, которые имелись в изобилии, обеспечивали нужды растений. Следовательно, хлорофилла самого по себе было достаточно. Однако почему же не развился пигмент, способный непосредственно поглощать часть спектра с наибольшей энергией фотонов? Ответ, вероятно, заключается в том, что химическое связывание элементов в молекулы имеет свои пределы. Просто число молекулярных комбинаций, которые могут производить ту же работу, что и хлорофилл, не бесконечно и хлорофилл, быть может, был лучшим соединением в то время и в том месте. Когда океан стал глубже, ограничения, возникшие для системы растений, приспособленной к хлорофиллу, компенсировались развитием вспомогательных пигментов. Но важно отметить, что химия хлорофилла сохранилась; действительно, вспомогательные пигменты не способны осуществить всю последовательность взаимодействий молекул, необходимую для образования сахара и других веществ, требующихся тканям.
Дата добавления: 2019-10-17; просмотров: 985;