Объекты изучения и перспективы
Что такое Флористическая география
Объекты изучения и перспективы
·
Флористическая география - один из наиболее рано сформировавшихся разделов географии растений. Флористическая география решает две задачи глобального масштаба. Первая - изучение географического распространения ареалов отдельных видов, в других случаях - надвидовых или внутривидовых систематических единиц. Вторая - сравнительное изучение флор, дислоцированных на континентах мира и в мировой акватории. Изучение флор отдельных территорий дает материал для флористического районирования, выделения на суше Земли флористических царств и более дробных флористических пространственных единиц: областей, провинций, округов и т. д. В результате создается флористическая иерархическая система, т. е. система соподчиненных единиц флоры - хорионов от флористического царства до района флоры.
Наряду с общими флористическая география решает и местные, региональные задачи, имеющие важнейшее народнохозяйственное значение. Среди указанных направлений отметим изучение закономерности структуры растительного покрова, распространения и распределения видов в региональном аспекте, в частности закономерности видового состава растительности - степей, прерий, лугов, болот, лесов и др. Однако география растительности - объект внимания прежде всего экологической географии, изучающей также многие проблемы экологии растений, особенно фитоценологии. Таким образом, флористическая география не только тесно сопряжена, но и взаимопроникает в другие ботанико-географические науки.
Давно уже накопились факты, свидетельствующие, что в разных местностях земного шара встречаются особые комплексы растительности, хорошо различающиеся как по флористическому (видовому) составу, так и по структуре жизненных форм. С другой стороны, бросается в глаза, что одни группы растений характеризуются очень широким распространением, другие - весьма ограниченным. Так, пальмы широко встречаются в тропиках обоих полушарий (пантропическое распространение), хвойные породы широкой полосой леса окружают Северный полюс (циркумполярное распространение). Основные лесообразующие породы циркумполярной тайги - сосны, пихты, лиственницы, ели. Тайга канадская по составу жизненных форм и структуре леса в целом очень напоминает нашу сибирскую. Конечно, виды хвойных в Западном полушарии - одни, а в Восточном - другие. Близкородственные виды, сменяющие друг друга на разных пространствах Земли, называют замещающими, иначе - корреспондирующими, или викарирующими. Подробнее о замещающих видах и ареалах будет сказано в главе об ареалах.
В развитии флористической географии видная роль принадлежит русским ученым и деятелям науки стран территории бывшего СССР. Это и понятно: необъятные пространства страны с отчетливо выраженными почвенно-растительными зонами и поясностью в горах, ярко очерченными геологическими водосборными районами, все еще сохраняющими пышный растительный покров, стимулировали развитие науки. Ботаники-географы установили добрую традицию изучать растительность и флору не отвлеченно, а с перспективой ее рационального хозяйственного использования. В наши дни выдвигается еще одна задача первостепенного значения - изучение способов обогащения и охраны флоры.
Лучшим образцом комплексного изучения некоторых территорий царской России с целью сельскохозяйственного освоения была работа Переселенческого управления (1905 - 1914). Бок о бок, в едином комплексе, выполняя одну общую народнохозяйственную задачу, работали ботаники и гидрологи, зоологи и почвоведы, экономисты и климатологи. Результатом явились разносторонние характеристики изучаемых территорий, их природное районирование. Последнее представляло собой действительно прочную научную основу организации сельскохозяйственного производства.
Конечно, работы по дробному ботанико-географическому, а тем более общему природному районированию трудоемки и требуют не только больших усилий, но и времени. Порой даже невозможно предвидеть, в каких отраслях наук о природе и в какой сфере практической деятельности дробное флористическое районирование будет использовано. Однако уже сейчас совершенно ясно, что оно служит необходимой составной частью общего природного районирования, составляющего научную основу агроклиматического районирования страны. Неоценимо значение дробного природного районирования, вплоть до районирования флоры в разработке научных основ интродукции растений и прогнозировании ее эффекта. В этрй связи академик Н. Вавилов отмечал, что каждый вид растения - кандидат в интродуценты и его ближайшие родичи должны быть "положены на глобус", т. е. на карту природных районов флоры. При этом легко обнаружить экологическую пластичность - поливалентность интродуцента.
Ботанико-географическая школа России и стран на территории бывшего СССР характеризуется оригинальностью и широтой научно-теоретических обобщений. Русские ботаники-географы, экстраполируя данные, полученные на неохватных просторах страны, обоснованно приходили к широким обобщениям в масштабе всего Евразийского континента, особенно северной, внетропической его части. Здесь надо отметить целую серию выдающихся работ А. Краснова, Е. Вульфа, А. Ильинского, М. Попова и др.
Обобщающая работа А. Тахтаджяна "Флористические области Земли" появилась в 1978 г. В ней собрана научная информация, полученная при региональных ботанико-географических исследованиях. Основываясь на строго системном подходе, проанализированы интегральные свойства основного объекта исследований, каковым представляется естественноисторический вид во всех его связях, но главным образом его структура и динамика со средой обитания. В результате даны оригинальные характеристики шести флористических царств, 34 областей и 147 провинций. Система флористического районирования земного шара заслуживает дальнейшей разработки. Дело в том, что она, к сожалению, доведена только до провинций, т. е. лишь до третьей степени (царство - область - провинция). Решение конкретных задач, например природоохранных, требует более дробного районирования, доведенного до четвертой-пятой степени.
Различают геоботаническое районирование, флористическое и ботанико-географическое. Каждое из них преследует определенные цели, опирается на соответствующие методы исследования, но между ботанико-географическим и флористическим районированием есть много общего. Свидетельством указанному служит тот факт, что высшие таксоны ботанико-географического районирования - ботанико-географические области - во многом совпадают с флористическими царствами - высшими таксонами флористического районирования. Упомянутое явление легко объяснить тем, что ботанико-географические области выделяются на основе флорогенетического принципа, т. е. общности истории происхождения и развития той флористической основы, на базе которой формируются растительные сообщества. При выделении флористических царств важнейшее значение придается сочетанию систематических групп, эндемичным родам и семействам и другим показателям, тесно связанным или вытекающим из флорогенетических характеристик. Ботаники-географы нередко отождествляют понятия "флористическое царство" и "ботанико-географическая область".
Понятие об ареале
·
·
Слово "ареал" происходит от латинского слова аrеа - площадь, пространство. В биологической литературе под ареалом географическим (аrеа geographica) понимают пространство на поверхности Земли (или в акватории), занятое каким-либо видом растения (или животного). В пределах ареала вид присутствует в местообитаниях, соответствующих его экологии. Можно говорить об ареале не только отдельных видов, но и более крупных систематических групп (таксонов), их объединяющих. Можно вычерчивать ареалы растительных группировок, например площадей распространения степей, разных типов лесов, лугов, пустынь и т. д. Учение об ареалах, иначе ареология, ставит своей задачей изучить области распространения (ареалы) конкретных систематических единиц флоры или растительности. Данную науку в последнее время именуют также хорологией.
В результате изучения ареала какой-либо систематической единицы в природе, по литературным данным или гербарным материалам его вычерчивают на географической карте. Техника изображения ареала следующая:
на карте страны или другой территории местонахождение организмов изучаемого вида обводят контурной линией, а затем выделенную территорию окрашивают или заштриховывают;
все известные местонахождения особи изучаемого вида отмечают точками, кружками или какими-либо другими условными знаками, затем крайние пункты местонахождения соединяют сплошной линией (или не соединяют). Первый метод - контурный - чаще применяют при изображении крупных таксонов на больших площадях распространения, второй способ - точечный - при детальных флористических исследованиях.
В природе невозможно найти два идентичных ареала; они обязательно различаются размерами, формой, характером границ. Форма ареала зависит от ряда причин. К важнейшим из них относят исторические судьбы данного вида, разнообразие (пестроту) местообитаний, соответствующих данному виду, взаимоотношения с другими видами растений и животных (включая человека). Различают ареалы природные, т. е. сформировавшиеся естественным путем, и искусственные. Последние возникают под влиянием хозяйственной деятельности человека. Размеры ареала чрезвычайно разнообразны, что для каждого таксона обусловлено исторически. Выяснение истории изучаемого вида, рода или семейства во времени и пространстве составляет главную цель исторической географии.
Пределы распространения вида, его границы определяются комплексом причин. К числу непреодолимых препятствий для распространения вида относят механические: горные хребты, водные пространства, пустыни. Климат также может представлять непреодолимый рубеж. Долгое время, вплоть до 19 столетия, считалось, что климатические границы ареала обусловлены только температурами. В наши дни выяснено, что климатическую границу ареала определяет совместное влияние двух факторов - температуры и влажности. Строго говоря, граница ареала зависит от совокупности факторов, важнейшие из которых, помимо климатических, - почвенно-грунтовые условия и биотические (конкуренция с другими растениями).
Иногда выделяют исторические границы ареала, которые сформировались в отдаленные геологические периоды и не могут быть объяснены современными причинами. Границы ареала, определяемые экологическими факторами, в том числе и биотическими (например, наличие конкурентных видов или отсутствие симбиотического компонента), как правило, подвижны. В одних случаях границы ареала расширяются, в других сокращаются. В первом случае имеют место прогрессирующие, т. е. растущие, ареалы, во втором - редуцирующиеся, сокращающиеся. Изучение изменений границ в исторической перспективе - одна из первоочередных задач исторической географии.
Границы ареала на всей протяженности могут определяться разными факторами. На одних отрезках границы, например южной, может быть дефицит влаги или засоленность почвы; на северных - дефицит тепла, а на западном или восточном участке продвижение вида ограничивает эколого-ценотическая обстановка. Таким образом, ограничивающая (экстремальная) обстановка на разных участках границы ареала обусловлена, как правило, разными факторами. Следовательно, и популяции вида, формирующиеся под влиянием экстремальной обстановки, будут качественно неодинаковы. В разных направлениях будет развиваться и процесс адаптации - микроэволюции. На границах ареала дислоцированы наиболее контрастные популяции как по структурно-физиологическим, биохимическим, так и по хозяйственно ценным свойствам. Здесь разнообразнее и материал для работы селекционера по созданию сортов.
Изложенное представление вполне согласуется с дарвиновской миграционной концепцией. По Ч. Дарвину, вид, возникнув где-либо, начинает затем расселяться в разных направлениях потенциально по всем радиусам. На пути расселения популяции вида там и здесь сталкиваются с новыми экологическими условиями, в том числе и биоценотическими, и вступают с ними в определенные взаимоотношения, нередко антагонистические: по отношению к компонентам (видам) фитоценозов - конкурентные. Указанные взаимоотношения и вызывают процесс адаптивных изменений - микроэволюцию.
Величина ареала зависит не только от названных объективных причин. Нередко важную роль при очерчивании границ ареала играют субъективные взгляды исследователя, его отношение к пониманию биологического вида. Суть в том, что одни ученые - флористы и систематики понимают вид крупно, в масштабе линнеевских видов - линнеонов; другие - узко и конкретно, в масштабе жорданонов. Жорданон - внутривидовой таксон, введенный Г. Дефризом и названный в честь французского ботаника А. Жордана, известного тем, что в рамках одного вида растения семейства Капустные (Brasstcaceae), а именно крупки весенней (современное наименование веснянка весенняя - Erophila verna), описал 200 мелких наследственных форм, которые предложил рассматривать в ранге видов.
Диапазон ареалов по величине очень широк. Известны виды, распространенные на всех континентах мира, кроме Антарктиды. Их ареал простирается почти на половину суши нашей планеты. Данные виды называют космополитами (от греч. kosmos - вселенная и polites - гражданин). Термин ввел в ботаническую географию А. Декандоль (1855). Он же привел список растений-космополитов.
К космополитам относят некоторые водно-болотные растения, например тростник обыкновенный (Phragmites communis), рогоз широколистный (Typha latifolia) и рогоз узколистный (Г. angusufolia), род рясок (Lemna). Среди космополитов многие сорняки полей и рудеральные растения: пастушья сумка (Capsela bursa-pastorts), мятлик однолетний (Роа аппиа), марь белая (Chenopodium album), подорожник большой (Plantago major). К космополитам можно отнести род одуванчиков (Таrахасит), семейство Мятликовые (Роасеае) и др. Космополитные таксоны формируют соответственно и космополитные ареалы (рис. 1). Последние служат антиподами эндемичных ареалов.
Космополитизм в мире растений пропорционален рангу систематических групп. Так, значение соотношения космополитных и некосмополитных видов намного меньше, чем соотношения таковых родов. Отношение космополитных семейств к некосмополитным в процентах превосходит таковое у родовых групп.
Ареалы многих видов растений ничтожно малы. Рекордно миниатюрный ареал имеют замечательный среднеазиатский вид нидзведския семиреченская (Niedzwedzkia semiretschenskia) и сибирский вид мегадения Бардунова (Megadenia bardunovii). Ближайшие родичи упомянутых видов находятся в тропиках. В Крыму незначительный по площади ареал занимает сосна Станкевича (Pinus stankewiczii), а на Кавказе - сосна эльдарская (Р. eldarica).
Миниатюрные ареалы имеют, как правило, древние эндемичные виды - палеоэндемики. От последних следует отличать неоэндемики - молодые виды, еще не успевшие
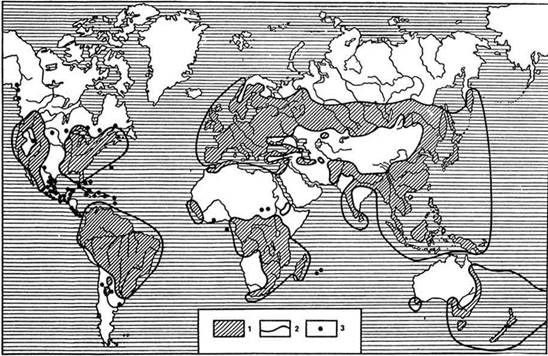
Рис. 1. Ареал папоротника орляка (Pteridium aquillinum) как пример космополитического распространения:
1 - пространства, где орляк встречается массово; 2 - граница массового распространения орляка; 3 - участки, где орляк встречается спорадически достаточно широко распространиться по земной поверхности, например некоторые виды колокольчика (Campanula), встречающиеся только на Кавказе.
Типы ареалов
Любой из существующих ареалов вида (рода, семейства) или растительной группировки имеет свой возраст, структуру и историю развития. Поскольку биологические особенности отдельных видов неодинаковы, различны и темпы развития их ареалов. Пространственное развитие, т. е. увеличение площади ареала, связано с ростом численности представителей вида, их реакцией на условия окружающей среды, заселением новых экологических ниш - местообитаний.
Большинство видов покрытосеменных сформировалось уже в верхнем мелу и палеогене, т. е. около 80...120 млн лет назад. Следовательно, ареалы видов и надвидовых таксонов этой группы растений претерпевали существенные преобразования в связи с изменением климата и других условий жизни. Сопоставляя признаки структуры видов растений и эколого-географическую их дислокацию, можно получать достоверные данные, во-первых, о направлении и ходе эволюции видов, во-вторых, об историческом развитии их ареалов.
В пределах ареала вид, как правило, никогда не заселяет всю площадь сплошь. Его представители встречаются лишь на тех участках, которые по экологической характеристике наиболее соответствуют природе вида. В этой связи возникло понятие о топографии вида, т. е. дислокации и частоте встречаемости его представителей в зависимости от экологических условий в пределах ареала: химизма почвы, ее влажности, взаимоотношений с другими видами фауны и флоры, рельефа местности и др.
Изложенное необходимо иметь в виду при определении двух типов ареалов - сплошных и разорванных, иначе - прерывистых, или дизъюнктивных (от лат. disjunctio - разобщение).
В сплошных ареалах особи вида обязательно заселяют свойственные ему местообитания на всем пространстве ареала. Так, виды клюквы (род Oxycoccus), как правило, присутствуют на всех сколько-нибудь хорошо развитых болотах северной лесной зоны. Напрасно бы мы искали клюкву в других местообитаниях, скажем, в пойменных лугах или дубравах. Однако даже в рамках сплошного ареала вследствие разных причин отдельные свойственные данному виду местообитания могут быть и не заселенными его представителями. Ярким и широко известным примером выборочной встречаемости вида в рамках огромного ареала служит тростник (Phragmttes communis). Его заросли можно найти лишь по берегам рек, озер и болот или на территориях с высоким стоянием грунтовых вод. В тех местах, где грунтовые воды находятся глубоко, например на водоразделах, тростника нет. Изучение распространения (топографии) популяций внутри ареала в зависимости от местообитаний - задача экологической географии. Наглядные примеры сплошных ареалов представляют виды сосна сибирская (Pinus sibirica) и ель сибирская (Picea obovata) (рис. 2).
В отличие от сплошных прерывистые ареалы характеризуются существенно большим пространственным разъединением отдельных заселенных участков, при котором связь между популяциями вида немыслима. Разобщенность отдельных участков столь велика, что воссоединение их при естественном распространении невозможно. Величина разъединенных участков бывает различна. Один из них может быть крупным, остальные мелкими. Разобщенность участков прерывистого ареала вызвана историческими причинами (вымирание вида в некоторых частях ареала, опускание участков суши, перемещение материков, влияние ледника и т. д.), а не деятельностью человека.
Согласно современным данным, виды возникают в какой-либо одной местности, поэтому первичный ареал должен быть сплошным, а разъединенный - явление вторичное.
Один из ярких примеров прерывистого ареала - ареал дерена шведского (Chamaepertclymenum suedicum). Ареал данного вида, приуроченного к приморским странам умеренного и арктического климата, состоит из двух четко обособленных участков: северо-запад Европы и северо-восток Северной Америки и дальний восток Азии и северо-западная часть Америки (рис. 3). Приведем три примера разъединений ареалов:
· первый - это арктоальпийские виды, представленные в Альпах, горах Средней и Южной Европы и в то же время в арктической области Старого и Нового Света: ива травянистая (Salix herbaced), ломонос альпийский (Clemaus alpina), смолевка бесстебельная (Silene acaulis), василисник альпийский (Thalictrum alpinum) и др.;
· второй - Европа и Северная Америка - виды, представленные участками ареалов, в Старом и Новом Свете: лобелия Дортмана (Lobelia dortmannii), осока желтая (Carex flava), плаун заливаемый (Lycopodium inundatum) и др.;
· третий - дизъюнкция Крым - Закавказье - Средняя Азия имеет общеизвестный вид: безвременник теневой (Colchicum umbrosum).
Многие виды имеют дизъюнкции "местного масштаба". Так, один из видов володушки, а именно володушка лютиковидная (Bupleurum ranunculoides), произрастает на трех разобщенных участках: на Урале, в Курской и Воронежской областях и в Карпатах.
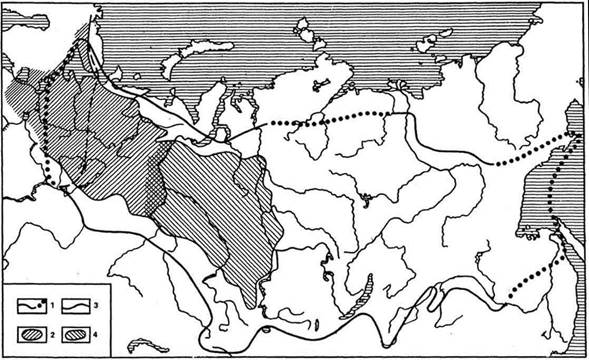
Рис. 2. Ареал ели сибирской (Picea obovata) как пример сплошного ареала:
1 - граница ареала; 2 - пространство, в пределах которого Р. obovata является основным лесообразователем зонального типа (в западной части этого пространства постепенно замещается Р. abies); 3 - восточная граница ареала Р. abies; 4 - область господства ели, пихты и сосны сибирской, на севере - ели и лиственницы в зональных типах леса
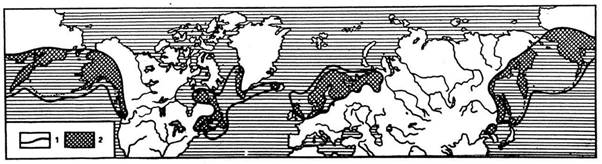
Рис. 3. Ареал дерена шведского (Chanmepericlymenum suedicum) как пример прерывистой дислокации вида, по А. Толмачеву:
1 - граница ареала; 2 - пространства суши, на которых произрастает дерен шведский
Дата добавления: 2017-02-20; просмотров: 915;