Ретикулярная формация, кора и парадоксальный сон.
В последние годы нейрофизиологи и нейропсихологи обратили внимание на особого рода состояние, периодически наступающее во время сна, которое по ряду признаков явно отличается от обычного сна и от бодрствования. Это состояние характеризуется быстрой электрической активностью ЭЭГ коры подобной в со-
стоянии бодрствования, сопровождается полным исчезновением мышечного тонуса, особенно шейных мышц и быстрыми движениями глазных яблок. Оно было впервые описано на людях в 1953—1955 гг. Азеринским и Клейтманом (73), в 1957 г. Дементом и Клейтманом (74), а у животных — Дементом в 1958 г. (75). Такая картина корковой электрической активности во время сна подробно исследована Жуве и сотр. в 1961—1965 гг. (13, 78, 79) и многими другими. Это своеобразное состояние сна известно под разными названиями, но в настоящее время наиболее принята терминология
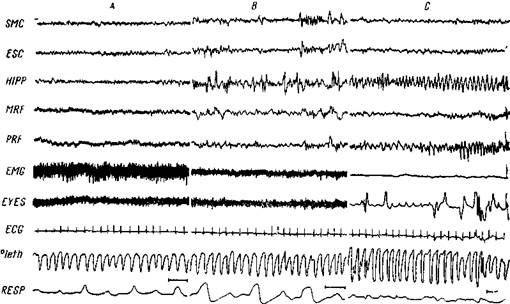
Рис. 107. Полиграфическая запись двух состояний сна
А — бодрствование: быстрая корковая и подкорковая активность В — медленный сон' корковые и подкорковые веретена и медленные волны Хорошо выраженная активность шейных мышц (EMG) Отсутствие движений глаз С — парадоксальный сон быстрая корковая активность такая же, как при бодрствовании Регулярная тета-активность в вентральном гиппокампе (HIPP) Фазическая активность в ретикулярной формации варолиева моста (PRF) Полное исчезновение электрической активности шейных мышц и быстрых движений глаз. Изменения дыхяния (RESP) и плетизмограммы (Pleth ). SMC — сенсомоторная кора, MRF — ретикулярная формация среднего мозга Калибровка 1 сек , 50 мкв (Жуве)
французских авторов — парадоксальный или ромб-энцефалический сон и американских авторов — стадия быстрых движений глаз.
Как известно, обычный сон, или, как его называют, медленный сон, в основном характеризуется двумя последовательными изменениями в электроэнцефалограмме: сначала появлением веретен, с частотой 15—18 в сек., а затем высоковольтной активностью — 2 — 4 в сек., распространяющейся по коре и подкорковым структурам. Эти две стадии сопровождаются тонической активностью шейных мышц и почти полным отсутствием движений глазных яблок.
Парадоксальный сон существенно отличается от медленного сна по электрографическим и внешним поведенческим проявлениям. Он обычно следует за медленным, длится в среднем 6—8 мин., периодически повторяется несколько раз в течение сна и в общей сложности составляет 20—25% всего медленного сна (Демент, 76, 77).
У кошек во время парадоксального сна в неокортексе регистрируется быстрая низковольтная активность — состояние десинхронизации, которая напоминает картину корковой активности при ярко выраженной тревоге или внимании. При этом в гиппокампе регистрируется регулярный тета-ритм (5—7 в сек.) (ряс. 107).
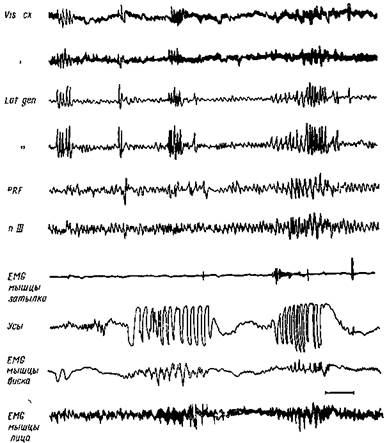
Рис. 108. Фааические феномены во время парадоксального сна
Нормальная кошка через 3 дня после энуклеации обоих глаз. Монофазные пики группируются в псевдоверетена в ретикулярной формации варолиева моста (PJRF), в ядре глазодвигательного нерв? (п III) в ядре наружного коленчатого тела (Lat gen ) и зрительной коре (Vis ex") Видны фазические подергивания усов, мышц лица и области виска, отсутствует электрическая активность шейных мышц (BMG). Калибровка 1 сек , 50 мкв (Жуве)
Наиболее ранним признаком парадоксального сна является возникновение монофазных длительных острых волн по 8—10 в сек. в ретикулярной формации моста и в латеральном коленчатом теле. Они на 30—90 сек. предшествуют движениям глазных яблок и быстрой биоэлектрической активности (рис. 108). Так как они не возникают после разрушения ретикулярного ядра в задней части моста или же ретикулярной формации ствола мозга перед мостом (Жуве, 80), то надо связать их происхождение с этими отделами мозга.
У человека, при парадоксальной стадии сна, также наблюдается характерная для бодрствования частая низковольтная ЭЭГ. Лишь частота альфаволн в затылочной доле на 1—2 гц меньше, чем при бодрствовании. Но у человека наиболее постоянным и специфическим отличием парадоксального сна считают появление вспышек острых треугольных волн по 2—3 в сек. в коре большого мозга, особенно сильно в ее центральной и теменной области. Каждая вспышка продолжается несколько секунд и предшествует или частично совпадает с быстрыми движениями глазных яблок (Жуве, 81). Во время парадоксального сна обычно наступают сновидения.
Парадоксальный сон характеризуется также особого рода поведенческими проявлениями. Во-первых, это — мышечная атония, особенно шейных
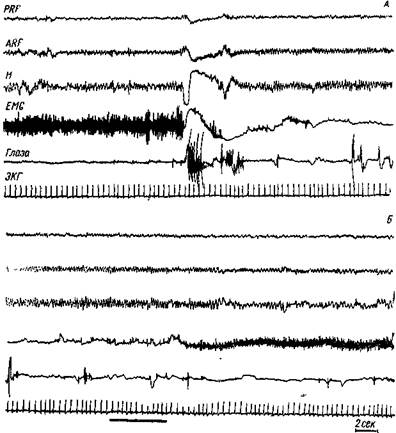
Рис. 109. Парадоксальный сон у полностью декортицированной кошки.
А—сначала поведение животного спокойное, но нет подкорковой медленной активности. Неожиданно голова падает (исчезает электрическая активность шейных мышц) и появляются быстрые движения глаз. На уровне дорзального гшшокампа (Я) и вентральной части восходящей ретикулярной формации (ARF)— уровень интерпедункулярного ядра— появляется медленная ритмическая активность 5 колебаний в сек. PRF — электроды в варолиевом мосту, на уровне ножек мозга. Запись Б — через 40 сек. после А. Черная линия внизу обозначает звуковой стимул большой интенсивности, животное просыпается. Калибровка 50 мне (Жуве)
мышц, с внезапным падением головы и полным расслаблением животного, которое обязательно принимает лежачее положение. При этом электрическая активность шейных мышц полностью исчезает. Во-вторых, быстрые движения глазных яблок (60—70 в мин.) при слегка открытой глазной щели и суженном зрачке. Менее часто наблюдаются у кошки быстрые подергивания ушей, челюстей, усов, губ, хвоста и конечностей (Демент, 75; Жуве, 78) (рис. 108).
Функциональное состояние ЦНС во время парадоксального сна меняется очень своеобразно. Установлено, что как спонтанная, так и вызванная активность корковых нейронов высокая, как при бодрствовании, и даже, может быть, выше (Росси, 82).
Очень характерно, что в опытах на людях условные сигналы, подкрепляемые болевым раздражением, вызывали во время парадоксального сна очень живые реакции пробуждения, а во время обычного медленного сна они не действовали (Вилльямс, 83). Значит, во время парадоксального сна избирательно повышается возбудимость в ЦНС, в частности, в коре большого мозга, в силу чего становится возможной условнорефлекторная деятельность. Отсюда надо полагать, что парадоксальный сон, при котором живот-
ное легко пробуждается, имеет важное биологическое значение. Он как бы является отражением периодической активации центральной нервной системы для подготовки животного к защите ог возможной опасности (Снайдер, 84).
Теперь установлено, что парадоксальный сон существенно не зависит от корковой деятельности, ибо он проявляется у декор!ицированны\ и хронических мезенцефалических животных (Жуве, 78) (рис. 109). По данным Жуве, пусковой центр парадоксального сна расположен в ретикулярной
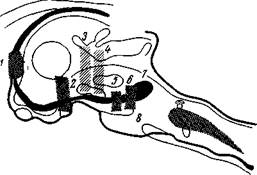
|
Рис. 110. Схема нервных ядер и i,\тей, участвующих в возникновении парадоксального сна
1 — перегородка (septum), 2 - субталамическая область (regio subtha. l^micus), 3 и 4 — разречы чсре,1 мезенцефатгическую ретикулярную формацию, 5— область ме/кду ножками мозга (regio mterpeduncularis), 6 — покрышка моста (tegmentum pontis), 7 — переднее ядро, s — заднее ядро ретикулярной формации моста (Наута и Кюйперс)
формации варолиевого моста и связан с корой головного мозга посредством специальной системы путей, отличных от восходящей активирующей системы. Это было установлено путем перерезок ствола мозга на разных уровнях.
Как видно из рис. 110, этот путь начинается от задней (8) и передней (7) частей ретикулярной формации моста, проходит через вентральную часть покрышки моста (6), область между ножками мозга (5), субталамическую область (2), перегородку (1) и поясную извилину, занимая, таким образом, лимбо-мезенцефалический круг (Наута, Кюйперс, 70).
Повреждение любого пункта этого круга устраняет проявление парадоксального сна, в частности, тета-ритм в гиппокампе и десинхронизацию ЭЭГ. Так на рис. 110 показаны разрезы 3 и 4 через мезенцефалическую ретикулярную формацию (восходящую активирующую систему), устраняющую десинхронизацию ЭЭГ при пробуждении, но не устраняющую десинхронпзацию во время парадоксального сна (Жуве, 80).
Парадоксальный сон можно вызвать искусственно электрическим раздражением не только ретикулярной формации моста, но и среднего мозга и гшшокампа и притом, как у интактных животных, так и у декортицированных. Более того, парадоксальный сон можно вызвать у интактных кошек раздражением кожных и мышечных нервов, если оно применяется в периоде медленного сна (Жуве и Жуве, 81), причем в этих случаях парадоксальный сон наступает со скрытым периодом 1—3 сек. Но у некоторых он наступает через 20—30 сек.
Характерно, что электрическим раздражением пускового механизма в варолиевом мосту нельзя вызвать парадоксальный сон подряд, друг за другом или после спонтанной фазы этого сна. Должно пройти по крайней мере 15—20 мин., чтобы новое раздражение вызвало парадоксальный сон. Необходимость такого длительного «рефракторного» периода указывает на участие нейрогуморального процесса. Во всяком случае установлено, что у кошки атропин (0,2 мг/к) подавляет парадоксальный сон, а эзерин усиливает его (Жуве и сотр., 85, 86). При внутривенном введении адреналина (1—5 мкгр/ /кг) никогда не наступал парадоксальный сон. А однократная инъекция большой дозы резерпина устраняет парадоксальный сон на много дней (Жуве, 85, 86). Все это дает нам право предположить, что каждый раз в период парадоксального сна в ЦНС продуцируется какое-то активное вещество, которое
-удерживается довольно долго и со своей стороны каким-то образом подавляет пусковой механизм парадоксального сна в ретикулярной формации варолиева моста. Вследствие этого парадоксальный сон может наступать только периодически.
Очень резкие количественные изменения претерпевает парадоксальный сон в процессе онтогенетического развития. Так, у новорожденных котят эта фаза занимает 80—90% всего времени и повторяется до семи раз в течение часа. С возрастом его общая продолжительность уменьшается до 25% и повторяется в течение часа только два раза (Валац, 87; Валац и Жуве, 88). Создается такое впечатление, что парадоксальный сон является онтогенетически более ранним явлением, чем медленный сон. Предполагается, что периодическая активация головного мозга, должно быть, имеет стимулирующее значение в онтогенетическом развитии ЦНС.
Специальное исследование парадоксального сна после искусственного лишения этого сна в течение многих часов и дней (когда человеку или животному — кошке — не дают возможности ложиться и так спать или будят во время наступления этой фазы), показало, что у кошки в последующем восстановительном периоде, когда она спала в нормальных условиях, процент парадоксального сна значительно увеличивался (Демент, 76; Жуве, 86). Это увеличение происходило в основном за счет укорочения периода медленного сна. Причем во время парадоксального сна в этом восстановительном периоде наблюдаются более интенсивные подергивания тела, хвоста, усов животного, напоминающие картину эпилептических припадков. Кроме того, у животных в этом периоде во время парадоксального сна повышена возбудимость слуховой системы, усилена пищевая потребность; они более беспокойны, более сексуальны. У людей в этом восстановительном периоде также наблюдалось компенсаторное увеличение парадоксального сна.
Все эти функциональные изменения свидетельствуют о том, что в моменты отдыха в пусковом механизме парадоксального сна, находящемся в ретикулярной формации варолиева моста, происходит спонтанное периодическое усиление активности, которая, вероятно, определенным образом связана с периодическим усилением определенного рода обмена веществ. Из активированного пускового механизма исходящая импульсация, должно быть, производит нейрогуморальное влияние на многие отделы головного мозга. В результате этого в одних нервных комплексах, в частности, в коре большого мозга и в двигательных центрах, возбудимость повышается, а в других, например, тонических и локомоторных координирующих аппаратах, она понижается. Поэтому в этом периоде в коре повышается способность к спонтанной деятельности, выражаясь, у людей в частности, в сновидениях, а подкорковые механизмы тонуса и локомоции лишаются способности к «спонтанной» или вызванной деятельности. И если во время парадоксального сна условные сигналы в состоянии вызвать адекватную реакцию, а другие раздражения вообще не действуют, так это должно быть происходит благодаря активированию хорошо развитых временных связей с двигательными центрами, в которых возбудимость была повышена.
Таким образом, активность коры большого мозга периодически поддерживается на некотором высоком уровне во время сонного состояния, благодаря периодическому активированию ретикулярной формации в области варолиева моста. Тем самым парадоксальный сон является очень важным защитным приспособлением для адекватного реагирования на жизненно важные внешние воздействия во время сонного состояния.
ЛИТЕРАТУРА
1. М о г u z z i G. a. M a g о u n H. W. EEG and Clin. Neurophysiol., 1949, 1, 455.
2. НарикашвилиС. П. Труды Ин-та физиол. им. Бериташвипи, 1950, 8, 135; 1953, 9, 133.
3. RhoinbergerM. В., Jasper Н. Н. Атег. т. Physiol., 1937, 119, 186.
4. MorisonK. S.,DempseyE. W. Amer. J. Physiol., 1942, 135, 289.
5. ЦкипуридзеЛ. Р. Труды Ин-та физиоп. им. Бериташвили, 1950, 8, 189 и 209.
6. S е g un d о J. P., A r a n a R. and F r e n с h J. D. J. Neurosurg., 1955, 12, 601.
7. L i n d s 1 e у D. В., В о w d e n J. W., a. M a g о u n H. W. EEG and Clin. Neurophysiol. 1949, 1, 475.
8. LindsleyD. В., SchreinerL. H., KnowlesW. B. a. Magoun H. W. EEG and Clin. Neurophysiol., 1950, 2, 483.
9. LinsleyD. B.,SchreincrL. H. a. Magoun H. W. J. Neurophysiol., 1949, 12, 197.
10. НарикашвилиС. П. и МониаваЭ. С. Вопр. нейрохирургии, 1962, № 1, 26.
11. В r e m e r F. С. R. Soc. Biol. Paris, 1935, 118, 1235.
12. Б р од а л А. Ретикулярная формация мозгового ствола. Анатомические данные и функциональные корреляции. М., Медгиз, 1960.
13. J о u v e t M. In: «The nature of sleep». Little, Brown a. Co., Boston, Mass., 1961, 188.
14. BatiniC., MoruzziG., PalestiniM., R о s s i G. F. a. ZanchettiA. Science, 1958, 128, 30; Arch. Ital. Biol., 1959, 97, 1.
15. MagnesJ.,MoruzziG. a. Pompeiano O. In: «The nature of sleep». Little, Brown a. Co., Boston, Mass., 1961, 57.
16. D e 11 P., В о n v a 1 1 e t M., et H u g e 1 i n A. In: «The nature of sleep». Little, Brown a. Co., Boston, 1961, 86.
17. НарикашвилиС. П. Неспецифические структуры головного мозга и воспринимающая функция коры больших полушарий. Тбилиси, 1962.
18. L i Ch. L. J. Physiol., 1956, 131, 115.
19. J u n g R. In: «Reticular formation of the brain». Little, Brown a. Co., Boston-Toronto, 1958. 423.
20. H a p и к а ш в и л и С. П. Успехи соврем, биологии, 1961, 52, 257.
21. С a j а 1 S. R a m о n у. Histologie du systeme nerveux de Гпогате et des vertebres. Paris, Maloine, 1909, 949.
22. M e t 11 e г F. A. J. Compar. Neurol., 1934, 61, 221, 509; 1935, 62, 263; 1935, 63, 25.
23. R о s s i G. F. а. В г о d a 1 A. J. Anat. (London), 1956, 90, 42.
24. Б e p и т о в И. С. Труды Ин-та физиол. им. Вериташвили, 1937, 3, 21.
25. М с С u 11 о с h W. S., G r a f C. a. M a g о u n H. W. J. Neurophysiol., 1946, 9, 127.
26. H u g e 1 i n A. et В о n v a 11 e t M. J. Physiol. (Paris), 1957, 49, 1171, 1201.
27. Dusser de BarenneJ. C. a. McCullochW. S. J. Neurophysiol., 1938,1, 69; 1941, 4, 287, 311.
28. БеритовИ. С.,БрегадзеА. Н. иЦкипуридзеЛ. Р. Труды Ин-та физиол. им. Бериташвили, 1943, 5, 241.
29. S 1 о a n N. a. J a s p e r H. EEG and Clin. Neurophysiol., 1950, 2, 317.
30. К a a d a R. Acta Physiol. Scand., 1951, 24, Suppl., 83, 1.
31. В r e m e r F. et T e r z u о 1 о С. Arch. Int. Physiol., 1952, 60, 288; 1953, 61, 86; 1954, 62, 157.
32. P о и т б а к А. И. и Б у т х у з и С. М. Докл. АН СССР, 1961, 139, 1502.
33. French J. D.,Hernandez-PeonR. a. LivingstonR. В. J. Neurophysiol., 1955, 18, 74.
34. J о u v e t M. et M i с h e 1 F. C. R. Soc. Biol. Paris, 1958, 152, 1167.
35. Серков Ф. Н.,МакулькинР. Ф. иРусевВ. В. Физиол. журн. СССР, 1960, 46, 408.
36. MoruzziG. In: «Brain mechanisms and consciousness». Springfield, 1954, 21; XV Int. Physiol. Congr., Abst. Rev., 1956, 269.
37. H a n b e r у J. a. J a s p e r H. J. Neurophysiol., 1953, 76, 252.
38. HanberyJ.,Ajmone-MarsanC.,Dilworth M. EEG and Clin. Neurophysiol., 1954, 6, 103.
39. VerzeanoM.,LindsleyD. A.,Magoun H. W. J. Neurophysiol., 1953,16,183.
40. H a p и к а ш в и л и С. П. Труды Ин-та физиол. АН ГССР, 1958, 11, 269.
41. J a s p e r H., N a q u e t R., К i n g E. E. EEG and Clin. Neurophysiol., 1955, 2, 99.
42. J a s p e r H. EEG and Clin. Neurophysiol., 1949, 1, 405.
43. P о и т б а к А. И. Труды Ин-та физиол. АН ГССР, 1958, II, 121.
44. Jasper H. H.,Ajmone-Marsan C. Res. Publ. Ass. nerv, ment., 1952, 30, 493.
45. H a p и к а ш в и л и С. П., М о н и а в а Э. С. Сообщ. АН ГССР, 1957, 19, 349.
46. A k i m о t о J., S a i t о A., N a k a m u r a J. In: «Neurophysiologie und Psychophysik des visuellen Systems». Springer 1961, 363.
47. S p enc er W. А., В г о о kh a r t J. M. J. Neurophysiol., 1961, 24, 26.
48. H e s s W. R. In: «Brain mechanisms and consciousness.» Oxford. Springfield, 1954,117.
49. H 6 s 1 i L., M о n n i e r M. Pfl. Arch. ges. Physiol., 1962, 275, 439.
50. РойтбакА. и ЭриставиН. Журн. высш. нервн. деят., 1965, 15, 971.
51. L e v у Ch. К. а. С h e r v i n P. Fed. Proc., 1960, 19, 288.
52. S p i e g e 1 E. A. Arbeit. Neurol. Inst. Univ. Vienna. 1919, 22, 418.
53. G 1 e e s P. a. W a 11 P. D. Brain, 1946, 69, 195.
54. M с L a r d у Т. Brain, 1948, 71, 290.
-55. С о w a n W. M. a. P о w e 11 T. P. S. J. Neurol. Neurosurg. Psychiatr., 1955, 18, 166.
56. 3 а г e p О. Межуточный мозг. Изд. Акад, Рум. Нар. Респ. 1962.
57. С о х о W. S. a. S n i d е г R. S. Fed. Proc., 1956, 15, 42,
58. W h i t e s i d e I. A. a. S n i d e r R. S. J Neurophysiol., 1953, 16, 397.
59. Б у т х у з и С. М. Сообщ. АН ГССР, 1962, 614.
60. Б у т х у з иС. М. Физиол. журн. СССР, 1965, 51, 47; Журн. высш. нервн. деят., 1965, 15, 1036.
61. Бехтерев В. М. Основы учения о функции мозга. 1905.
62. MettlerF. R.,AdesH. W.,LipmanE. a. CullerE. A. Arch. Neurol. Psychiatr., 1939, 41, 984.
63. A k e r t К. М., a. A n d e г s о n B. Acta Pliysiol. Scand., 1951, 22.
64. F о r m a n D. a. W а г d J. W. J. Neurophysiol, 1953, 20, 230.
65. S h i m a m о t о T. a. V e r z e a n о M. J. Neurophjsiol., 1954, 17, 278.
66. KoizumiK.,UshiyamaJ. a. BoroksC. McC. Amer. J. Physiol., 1958, 195, 1.
67. Б у т х у з и С. М. Сообщ. АН ГССР, 1961, 27, 613; 1962, 28, 363.
68. Albe-FessardD. EEG and С 1 i n. Neurophysiol., 1959, 9, 370.
69. Horde Ch. A. a. M e t t 1 e r F. A. Anat. Record., 1953, 115, 324.
70. N a u t a W. J. H. and К u у p e r s H. G. In: «Reticular formation of the brain». Boston-Toronto, 1958, 1.
71. H a p и к а ш в и л и С. П., Б у т х у з и С. М., и М о н и а в а Э. С. В сб. «Соврем, проблемы деятельности и строения ЦНС» Тбилиси, изд-во «Мецниереба», 1965, 39.
72. НарикашвилиС. П., БутхузиС. М. иМониаваЭ. С. Физиол. журн. СССР, 1960, 46, 653.
73. AserinskyE. a. Kleitman N. Science, 1953, 116, 273; J. Appl. Physiol.,, 1955, 8.
74. D e m e n t W. С. а. К 1 e i t m a n N. J. Exp. Psychol., 1957, 53, 339.
75. D e m e n t W. C. EEG and Clin. Neurophysiol., 1958, 10, 291.
76. D e m e n t W. C. Amer. J. Psychiatry, 1965, 122, 4.
77. 0 e m e n t W. C. Aspects anatomo-lunctionale de la physiologie de sommeil. Extrait Discussion generale. Lion, 1965.
78. J о u v e t M. Progress in Brain Research. Amsterdam. 1963, 1, 406.
79. J о u v e t M. et J о u v e t D. Psychiatric animal. E. Editor. Deselee de Brouwer, 1964.
80. J о u v e t M. Progress in Brain Research, 1965. 18, 19.
81. J о u v e t M. a. J о u v e t D. EEG and Clin. Neurophysiol., 1963, Suppl., 24, 133.
82. R о s s i G. F., F a v a 1 e E., H a r а Т., G i u s s a n i H., a. S а с с о G. Arch. Ital. Biol., 1961, 99, 270.
83. W i 1 1 i a m s H. L., H a n n m а с k J. Т., D a 1 у R. L., D о m en t W. C. a. L i i b i n A. EEG and Clin. Neurophysiol., 1964, 16, 269.
84. S n у d e r F. Amer. J. Psychiatry, 1965, 22, 4.
85. J о u v e t M. XXIII. Int. Congr, of Physiol. Sciences. Tokyo, Symp. VI. Neural mechanisms of conditioned reflex and behaviour, 1965, 344.
86. JouvetM.,VimontP.,DelermeJ. F. aJouvetM. C. R. Soc. Biol., 1964, 158, 756.
87. V a 1 a t z J. L. Ontogenese des differents etats de sommeil. Imprimerie des beaux-arts cammille annequin. Lion, 1963,
88. V a 1 a t z J. L., J о u v e t D. a. J о u v e t M. EEG and Clm. Neurophysiol., 1964, 17, 218.
Дата добавления: 2017-01-13; просмотров: 894;