Взаимодействие коры, ретикулярной формации и мозжечка.
Подробно изучено влияние раздражения мозжечка на спонтанную и вызванную активность отдельных нейронов ретикулярной формации (Моруцци, 36). Известно также, что многие эффекты раздражения мозжечка в основном осуществляются через ретикулярную формацию, хотя известны и пути экстраретикулярного характера. Однако относительно обратного влияния ретикулярной формации на мозжечок, несмотря на наличие хорошо изученных эфферентных связей, известно очень мало. Выполненные за последнее время работы (Леви и Червин, 51; Нарикашвили и Мониава, 10) показывают, что восходящее из ретикулярной формации влияние на мозжечок такого же характера, как и на кору больших полушарий (рис. 106.) Ото выражается в том, что при ретикулярном раздражении ответы мозжечка (на звук, свет и электрическое раздражение зрительной хиазмы) угнетаются или облегчаются, так же как и ответы коры больших полушарий, хотя и в меньшей степени и на менее короткое время. Таким образом, судя по эффекту облегчения, восходящее ретикулярное влияние одинаково сказывается на деятельность нейронов как коры больших полушарий, так и мозжечка.
Как видно из осциллографических исследований, восходящее ретикулярное влияние на кору большого мозга бывает как облегчающим, так и тормозящим. Если это сопоставить с некоторыми морфологическими данными об окончаниях волокон восходящей неспецифической системы вдоль по всей
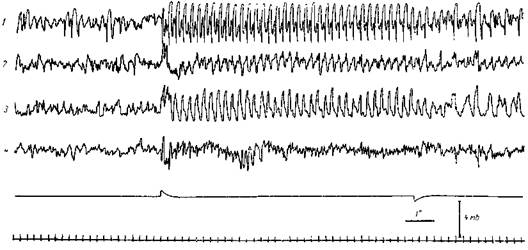
Рис. 106. Одновременное облегчение ответов коры больших полушарий и мозжечка под влиянием раздражения ретикулярной формации
Ненаркотизированная кураризованная кошка. Сверху вниз регистрируются потенциалы: задней супрасильвиевой извилины (1), средней латеральной извилины (2), задней латеральной извилины (3) и коры мозжечка (4). Нижняя сигнальная линия отмечает одиночные электрические раздражения (Зв, 0,5 мсек) зрительной хиазмы. Верхняя сигнальная линия отмечает электрическое раздражение мезенцефалической ретикулярной формации (Зв, 0,5 мсек, 200 в сек.). Хорошо видно значительное и одновременное облегчение ответов (возрастание амплитуды и регулярное возникновение) в зрительной коре и мозжечке при добавлении ретикулярного раздражения. В отличие от ответов зрительной коры, ответы моз/кечка облегчаются меньше и на менее короткое время (Нарикашвили и Мониава)
длине верхушечных дендритов в верхних слоях коры, то этому сложному явлению можно дать такое объяснение. Как это видно на рис. 9—II, неспецифические волокна подходят к основанию верхушечного дендрита, а затем тянутся по всей длине до первого слоя, касаясь лишь боковыми синапсами. Мы можем предположить, что когда при возбуждении этих волокон эффект ограничивается в основном активацией боковых синапсов, дендритный постсинаптический потенциал настолько будет слаб, что он своим электротоническим действием может повлиять на клетку облегчающим образом, деполяризуя ее постсинаптическую мембрану и аксонный холмик, не производя при этом анэлектротонической блокады пресинаптической мембраны. В таких случаях неспецифическая восходящая система окажет на кору облегчающее действие.
Но когда вовлекаются в реакцию и крупные терминальные синапсы на верхушечных дендритах первого слоя (см. рис. 30, 38), электротоническое действие дендритного тока может усилиться до такой степени, что оно произведет в пресинаптической мембране анэлектротоническую блокаду. Теперь неспецифическая система произведет торможение коры.
Дата добавления: 2017-01-13; просмотров: 731;