Об образовании нейронных систем, производящих образы внешнего мира.
Мы выше показали, что субъективные переживания в виде дифференцированных ощущений должны продуцироваться сенсорными звездчатыми клетками IV слоя (гл. II). Объединение корковых нейронов между собой осуществляется пирамидными нейронами, а также звездчатыми нейронами второго типа. Теперь перед нами стоит вопрос: каким образом сенсорные звездчатые и пирамидные ассоциационные нейроны осуществляют возникновение и репродукцию образов внешнего мира? По-видимому, первым здесь является вопрос о расположении афферентных волокон.
Афферентные таламические волокна, начинающиеся в специфических ядрах, оканчиваются преимущественно в IV слое первичных зон воспринимающих областей, но в некоторой мере в /// и V слоях тех же зон, а также во вторичных зонах, но везде они оканчиваются преимущественно на звездчатых сенсорных нейронах. Однако они оканчиваются в этих слоях и на малых пирамидных нейронах, аксоны которых не выходят из пределов коры большого мозга, т. е. на вставочных нейронах, а также на средних пирамидах /// и IV слоев, являющихся ассоциационными (Лоренте де Но, 14; О'Лири и Бишоп, 10; О'Лири, 32; Поляков, 11). Эти структурные отношения даны на рис. 61, где афферентные волокна из коленчатого тела оканчиваются на звездчатых клетках с короткими аксонами, а также на клетках Кахала с длинными аксонами, которые, как мы увидим ниже, служат для осуществления ориентировочных движений глаз.
Афферентные волокна, начинающиеся в ретикулярной формации головного мозга, должно быть, оканчиваются во всех участках коры большого мозга, но особенно много их в верхних слоях коры (Шайбель и Шайбель, 33). Посредством этих так называемых неспецифических волокон активируется вся кора (Лоренте де Но, 14; Демпси и Морисон, 34; Моруцци и Мэгун, 35; Джаспер, 36).
По Джасперу, раздражение неспецифических областей таламуса вызывает электрические эффекты во всех слоях коры, в то время как при раздражении специфических таламических волокон и ядер электрические эффекты отводятся только из IV и /// слоев.
Известно, что при раздражении одной части восходящей таламической системы или ретикулярных структур таламуса наступает общее торможение коры или отдельных ее областей. Эта функциональная особенность данной части таламической афферентной системы, по нашему мнению, находит свое структурное выражение в том, что данные афферентные волокна неспецифической таламической системы оканчиваются на апикальных дендритах в / и // слоях.
Раздражение определенной части неспецифических волокон или ретикулярной формации мозга производит общее облегчение в виде десинхронизации электрической активности с ослаблением медленных и усилением быстрых колебаний. Видимо, последнее происходит от того, что эти волокна оканчиваются на звездчатых нейронах, а также на соме вставочных и ассоциационных пирамидных нейронов IV и /// слоев коры, участвующих в образовании нервных кругов.
В последнее время стало известно, что в ретикулярной формации возбуждение может распространяться более или менее избирательно. Это проявляется в том, что при раздражении любого рецептора афферентная импуль-
сация по непрямому пути, через ретикулярные структуры таламуса, направлена главным образом в тот участок соответствующего анализатора, куда поступают афферентные импульсы от данного рецептора по специфическому пути.
Кроме того, путем локального электрического раздражения ретикулярных структур таламуса установлено, что разные участки их преимущественно влияют на определенные части коры (Хэнбери и Джаспер, 7). При повторных и более сильных раздражениях возбуждение может распространиться из данного участка, связанного в основном с определенным анализатором, на все сетевидное образование таламуса и тем самым повлиять и на другие анализаторы (Ройтбак, 37).
Афферентные таламические волокна специфической системы не оканчиваются непосредственно в коре на клетках проекционных пирамидных нейронов V и VI слоев. Они связываются с ними посредством вставочных пирамидных нейронов или звездчатых клеток второго типа. С проекционными же пирамидами отдаленных участков коры они связываются более сложным путем: посредством ряда вставочных и ассоциационных нейронов.
Вторым вопросом организации нейронных систем является вопрос о расположении и распространении аксонов вставочных и ассоциационных нейронов. Ассоциационные волокна, переходящие из одного участка в другой, оканчиваются в основном во //—/// слоях и лишь в меньшей степени в других слоях. По-видимому, они здесь оканчиваются на звездчатых нейронах второго порядка и на вставочных пирамидных нейронах. По Полякову (38), концевые разветвления аксонов ассоциационных нейронов в ///—// слоях существенно отличаются от оканчивающихся в нижних слоях; в верхних слоях они сильно ветвятся и каждая ветвь образует чрезвычайно тонкие и густые сплетения, окутывающие тела и дендриты расположенных здесь пирамидных и передаточных звездчатых нейронов. Эти ассоциационные нейроны, видимо, служат объединению деятельности сенсорных нейронов различных воспринимающих областей для создания образов таких объектов, которые воспринимаются через разные рецепторы.
Двусторонние связи между более или менее отдаленными участками коры проходят не только по корковым слоям, т. е. транскортикально, путем вставочных нейронов, но главным образом через подкорковое белое вещество при посредстве аксонов, ассоциационных нейронов.
Можно утверждать, что все малые и средние пирамидные нейроны с нисходящими или восходящими аксонами, а также передаточные звездчатые клетки с отходящими горизонтальными и вертикальными аксонами образуют замкнутые круги, где каждое клеточное звено связано с остальными звеньями непосредственно или при помощи нескольких других клеток. В каждом данном участке коры эти нейроны образуют замкнутые круги как в вертикальном направлении, создавая между клетками разных слоев двусторонние связи, так и в горизонтальном направлении между разными участками коры.
Эти нервные круги в виде ассоциационных двусторонних связей образуются в верхних слоях коры — //, /// и IV. Капперс, Хубер и Кросби (39) приводят многочисленные доказательства, что именно эти слои являются как высшим рецепторным, так и высшим интегративным субстратом. Особенно большое значение имеют в этом отношении // и /// слои, поскольку здесь больше всего малых и средних пирамидных нейронов с нисходящими и восходящими аксонами. Характерно, что в ядерной зоне зрительного анализатора аксоны многих пирамидных нейронов проходят через IV слой без отдачи коллатералей этому слою (О'Лири, 32). Это указывает на то, что в IV слое превалирует воспринимающая, сенсорная функция вследствие господства в нем сенсорных звездчатых клеток и окончаний афферентных таламических волокон.
Объединение более или менее сложных нервных комплексов предполагает наличие определенного объединяющего центра. Для объединения сен-
сорных элементов одной и той же воспринимающей области этот центр, должно быть, находится на периферии ее. Так, например, для зрительных звездчатых клеток он будет в 18 и 19 полях. Для объединения же сенсорных элементов разных областей такой объединяющий центр будет между ними в так называемых ассоциационных полях, теменных, лобных и височных. Это предположение высказывалось раньше (Эббеке, 40; Гольдштейн, 41; Блуменау, 42; Беритов, 43). В настоящее время оно также разделяется многими учеными (Пенфилд, Джаспер и др.). Наблюдения Пенфилда (42), показывающие, что раздражением височных долей у человека можно вызвать слуховые и зрительные воспоминания из прошлой жизни, лучше всего свидетельствуют о том, что височная область является главным ассоциационным полем для слуховых и зрительных восприятий. Гистологическое обоснование этих представлений мы находим в работах Бехтерева (9), Флексига (45), а в последнее время Полякова (46) по онтогенезу коры большого мозга человека.
Способ объединения сенсорных нейронов ядерной зоны одной воспринимающей области схематически представлен на рис. 65, где объединение дается в отношении зрительной области. На рис. 66 представлена схема объединения сенсорных нейронов двух разных областей — зрительной и слуховой. Однако сами сенсорные звездчатые нейроны, аксоны которых ветвятся около собственных клеток, не могут входить как самостоятельные промежуточные звенья в нервные круги. Эти клетки должны активироваться из нервных кругов или непосредственно через их коллатерали, или посредством вставочных нейронов, но аксоны этих звездчатых клеток не могут активировать данные круги. При наличии такой структуры система сенсорных нейронов, как уже указывалось, не всегда в состоянии вызывать непосредственно то или другое внешнее действие, например, ориентировочную реакцию. Но если при восприятии внешнего мира возникает ориентировочная реакция, то это происходит благодаря тому, что зрительные афферентные волокна одновременно с возбуждением сенсорных элементов активируют пусковой механизм ориентировочной реакции, как это изображено на рис. 61. Точно так же надо полагать, что при репродукции образа те нервные круги, которые активируют соответствующий комплекс сенсорных элементов, одновременно активируют в коре соответствующий пусковой механизм ориентировочной реакции.
Физиологические наблюдения также свидетельствуют о том, что сенсорные нервные элементы не должны обязательно входить как звено в замкнутые нервные круги. Как известно, через кору большого мозга осуществляются условные рефлексы и на такие раздражения, которые субъективно не ощущаются, как, например, неощущаемые раздражения рецепторов мышц, сухожилий и внутренних органов, а также на подпороговые сенсорные экстероцептивные раздражения (Гершуни, 47). Во всех этих условных рефлексах в образовании нервных связей или нервных кругов обязательно участвуют пирамидные нейроны и передаточные звездчатые нейроны без вовлечения сенсорных звездчатых нейронов. Можно утверждать, что при образовании условных рефлексов на субъективно ощущаемые раздражения временные круги могут возникать без прямого включения воспринимающих это раздражение сенсорных звездчатых клеток, как отдельного звена этих кругов. С этим предположением хорошо согласуется, между прочим, известный факт, что после контузии животного (голубя, кошки, собаки) ударной воздушной волной некоторой умеренной силы психонервная деятельность (например, в виде способности к запоминанию местонахождения пищи) исчезает, в то время как выработанное до контузии автоматизированное пищевое поведение, протекающее по принципу условного цепного рефлекса, сохраняется без изменения (Беритов, 48; Брегадзе, 49). Это, очевидно, объясняется тем, что от взрывной волны, как вообще от многих других повреждающих агентов, раньше и сильнее всего страдают наиболее высоко развитые сенсорные элементы. И если бы эти сенсорные элементы входили в нервные круги как их отдельные звенья, то условнорефлекторная деятельность должна была бы
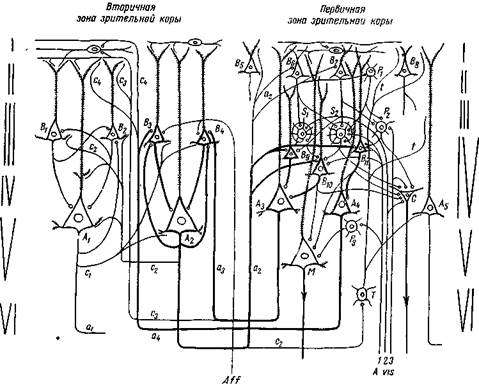
Рис. 65. Схема нервных кругов зрительной области, производящих зрительное восприятие и ориентировочную реакцию
S, — S2 — сенсорные звездчатые нейроны; Р,, Рг, Р, — передаточные звездчатые нейроны; Bt — Вг — вставочные пирамидные нейроны; А, — А4 — ассоциационные нейроны; М — пирамидный проекционный нейрон Мейнерта, С — проекционный глазодвигательный нейрон — звездчатая клетка Кахала; Т — вставочный нейрон Мартинотти; аа — а4 — аксоны ассоциационных нейронов; сэ — с4 — коллатерали их.
В Первичной зоне афферентные зрительные волокна (A. vis.) оканчиваются на клетках звездчатых и вставочных нейронов IV слоя, а также на клетке глазодвигательного нейрона.
Эти звездчатые и вставочные нейроны связаны с ассоциационными нейронами (А3 и А4) и с клеткой Мейнерта (М). С последней клеткой связано также таламическое волокно через звездчатую клетку Р3. Эта система нейронов при возбуждении зрительных таламических волокон производит, с одной стороны, восприятие объекта, а с другой — ориентировочную реакцию головы и глаз, направленную на фиксирование глаз на этот объект.
Аксоны от ассоциационных нейрснов первичней зоны А3 и А4переходятво вторичную зону зрительной коры и здесь заканчиваются на телах вставочных нейронов В„ и В4, которые своими аксонами конвергируют на ассоциационном нейроне А2, аксон которого а2 переходит в первичную зону и здесь оканчивается на клетках вставочных нейронов IV слоя (В,, Вю, Вц). Так создаются замкнутые нейронные круги. Они обозначены толстыми линиями. Эти круги активируются не только от одновременного возбуждения всех трех зрительных путей — 1, 2, з, но и от каждого из них в отдельности
Ассоциационные волокна, переходящие из вторичной зоны, оканчиваются также на вставочных пирамидных и звездчатых передаточных клетках II слоя.которые со своей стороны связаны с сенсорными звездчатыми клетками. При зрительном восприятии все эти нервные связи также активируются и тем создается функционально связанная система, которая сохраняется на более или менее длительное время и служит для репродукции образов всего объекта при воздействии только одной части его на глаз. Коллатераль с2 аксона ассоциационного нейрона А2 оканчивается в первичной зоне на вставочной клетке Т, аксон которой касается дендритов в верхних слоях, в частности, нейрона АЕ. Точно так же аксоны ассоциациоиных нейронов А3 и А4, перейдя во вторичную зону, отдают коллатерали сэ и с4, оканчивающиеся главным образом на дендритах верхних слоев Через эти коллатерали происходит активация дендритов соседних вставочных и ассоциационных нейронов и тем самым происходит общее торможение их. Это обеспечивает локализацию возбуждения в данной функциональной системе. На схеме дается связь с неспецифическими таламическими волокнами (А//) вторичной зрительной зоны
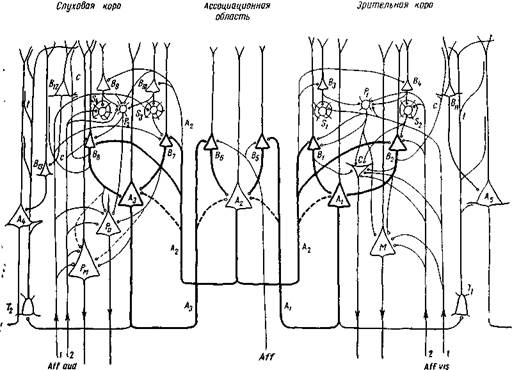
Рис. 66. Схема нервных кругов, объединяющих слуховую и зрительную воспринимающие области коры для восприятия и создания образа звучащего объекта
Буквенные обозначения те же, что на рис.65. От воздействия данного объекта афферентные волокна Aff. aud. и Aff. vis. в соответствующих воспринимающих корковых областях активируют сенсорные звездчатые нейроны IV слоя (S, — S2), передаточные звездчатые нейроны Pt и Р2, а также вставочные нейроны (В, — Вг, В, — В,). Через эти нейроны активируются ассоциационные нейроны (А, и Аз), аксоны которых уходят в ассоциационное полей конвергируют на вставочных нейронах В,—В6. Последние активируют ассоциационный нейрон А2, аксон которого уходит в слуховую и зрительную воспринимающие области и здесь оканчивается на вставочных нейронах IV слоя. Так образуются замкнутые круги между слуховыми и зрительными зонами. Эти круги\ обозначены толстыми линиями. От ассоциационных нейронов этих кругов коллатерали поднимаются в' слой II, возбуждая здесь вставочные нейроны. Через эти же нейроны А2 дополнительно возбуждаются как сенсорные нейроны (S, — S4), так и проекционные нейроны Мейнерта (АО и Кахала (С). Активация всей этой системы нейронов производит восприятие и создает зрительно-слуховой образ данного объекта и вызывает ориентировочную реакцию Эта функциональная система удерживается на более или менее длительное время. Coiласно этой схеме, при воздействии какой-либо части объекта (на схеме соответствующие афференты переданы цифрами 1, 2 Aff. aud. и 1, 2 Aff. vis.) возбуждается вся означенная система нейронов с рспроздк» цией всего образа воспринятого объекта и с наступлением соответствующей ориентировочной реакции головы. Aff. — неспецифический афферентный путь в ассоциационной области Р0 — проекцш нный экстрапирамидный нейрон ориентировочной реакции. На клетке его оканчиваются коллатерали аксонов афферентных нейронов слуха и аксонов от вставочных нейронов IVслоя 1'м — проекционный экстрапирамидный нейрон для общих движений. Т, и Т2 — нейроны Мартинотти, аксоны которых разветвляются в верхних слоях (t) на дендритах, в частности, дендритах ассоциационных нейр<нсв А, и А5 и вставочных |нейронов В,,, В12, В13, производя общее торможение вокруг возбужденных нейронных кругов; с —коллатераль аксона вставочного нейрона из этих нервных кругов, оканчивчющ яся на дегдритах соседних нейронов для общего торможения
исчезнуть наравне с психонервной. Надо думать, что нервные круги связываются с сенсорными элементами или непосредственно при помощи своих коллатералей, или же через дополнительные вставочные нейроны.
Итак, сенсорные звездчатые нейроны объединяются посредством вставочных и ассоциационных пирамидных нейронов, а также посредством передаточных звездчатых нейронов, т. е. второго типа. Эти сенсорные нейроны должны активироваться коллатералями входящих в нервные круги пирамидных нейронов непосредственно или через дополнительные вставочные нейроны.
Но сенсорные нейроны с околоклеточной аксонной сетью не должны входить как звенья в эти круги.
Далее, можно утверждать, что дендриты пирамидных нейронов, которые снабжены шшшками, не должны участвовать в образовании нервных кругов, ибо, как уже указывалось выше, они проводят возбуждение с большим декрементом, которое до тела клетки не доходит.
Аксоны проекционных пирамидных нейронов также не должны участвовать в образовании этих корковых кругов, ибо их коллатерали, за исключением тех, которые без ветвления поднимаются к первому слою, не выходят из пределов соответствующего V слоя, причем заканчиваются они в основном на базальных дендритах пирамидных нейронов. Аксоны проекционных пирамидных нейронов, выходя из коры, редко дают коллатерали (Бехтерев, 9; Лоренте де Но, 14; О'Лири и Бишоп, 10).
Итак, проекционные пирамидные нейроны не должны участвовать в образовании нервных кругов при восприятии и создании образов внешнего мира. Они возбуждаются при афферентной импульсации по коллатералям из тех нервных кругов, которые образуют вставочные и ассоциационные пирамидные нейроны при этой импульсации (см. рис. 66).
Аксоны проекционных пирамидных нейронов оканчиваются в средних отделах головного мозга, нейроны которых со своей стороны оканчиваются в коре. Поэтому проекционные пирамидные нейроны должны участвовать в образовании нервных кругов, объединяющих кору и подкорковые отделы, прежде всего промежуточный мозг (таламус), как его специфические ядра, так и ретикулярную формацию. В этих нервных кругах, значит, экстрапирамидные пути и специфические и неспецифические афферентные таламические волокна составляют основные звенья.
Когда на сетчатку глаза (животного или человека) действует какой-либо объект, то возбуждаемые при этом зрительные волокна по прямому специфическому пути не только активируют определенный комплекс зрительных сенсорных звездчатых клеток, но и одновременно активируют пусковой механизм для фиксирования глаз на данный объект для лучшего его восприятия. Как указывалось выше, в 17 поле находится множество так называемых клеток Кахала, длинные аксоны которых заканчиваются в переднем двухолмии. На телах этих клеток контактируют афферентные таламические волокна из коленчатого тела как через звездчатые клетки, так л непосредственно (см. рис. 61). Мы считаем, что эти клетки вместе с пирамидными клетками Мейнерта составляют пусковой механизм ориентировочных движений глаз и головы. Можно сказать, что восприятие объекта и фиксирование глаз на него составляет единый, неразрывный процесс и это важное физиологическое явление очень хорошо выступает на схеме ШкольникЯррос (16), представленной на рис. 61.
Однако наличие замкнутых кругов не означает еще, что возбуждение какого-либо звена обязательно приведет к направленному распространению возбуждения по соответствующим кругам. Свободный переход возбуждения от одного звена данных кругов на другое вырабатывается в основном в индивидуальной жизни и происходит как обязательное явление при определенных условиях.
Мы выше показали, что целостность корковой деятельности каждый раз является результатом возбуждения определенных нервных комплексов и общего торможения всей остальной коры. При этом мы показали, что общее торможение простирается как на эфферентную пирамидную систему, обусловливающую внешние реакции, так и на промежуточные и ассоциационные нейроны, участвующие в производстве спонтанной электрической активности коры.
Все эти факты дают нам возможность утверждать, что каждый раз при репродукции доминирующего образа, направляющего поведение, наряду с возбуждением одних нервных комплексов, производящих данный образ, должно происходить общее торможение вокруг них, а в некоторой мере и во всей коре.
По-видимому, смена одного индивидуального поведения, направляемого образом, на другое, направляемое другим образом, обусловливается,
с одной стороны, возбуждением комплекса нервных элементов последующего образа и отвечающей ему ориентировочной реакцией головы, а с другой стороны, торможением нервных комплексов, участвующих в производстве предыдущего образа. Когда собака, согласно данному доминирующему образу, идет в определенное место и съедает там всю пищу или не находит ее там, у нее возникает новый образ отсутствия пищи в этом месте. Соответственно исчезает ориентировочная реакция к этому месту. Этим самым снимается общее торможение, вызванное предыдущим образом. Вследствие этого какой-нибудь другой образ местонахождения пищи может стать доминирующим и вызвать соответствующую ориентировочную реакцию головы и ватем направленное движение уже к этому месту пищи.
Итак, можно утверждать, что целостная интегративная деятельность коры большого мозга во время любого индивидуального поведения, направляемого образом местонахождения пищи, так же, как при поведении в ответ на Непосредственное восприятие этого места, выражается в преобладании возбуждения над торможением в определенных нервных комплексах и, наоборот, в преобладании торможения над возбужением во всех соседних и отдаленных нервных комплексах коры. В этом выражается целостная интегративная деятельность коры, в результате которой наступает определенное движение головы и последующий целенаправленный поведенческий акт.
ЛИТЕРАТУРА
1. Б е р и т о в И. С. и Г е д е в а н и Д. М. Сообщ. АН ГССР, 1941, 2, 157.
2. Б е р и т а ш в и л и И. С. Труды Ин-та физиол. АН ГССР, 1937, 3, 21.
3. ГедеванишвилиД. М. Труды Ин-та физиол. АН ГССР, 1943, 5, 39.
4. Dusserde BarrenneJ. G. ,McCullochW. S. Symp. dedicated to prof» Beritashvili. Tbilisi, 1936, 15.
5. С 1 а г k G., К а о L i a n g С h о u, G i 11 a s р у С. С. а. К 1 о t s D. A. J. Neurophysiol., 1949, 12, 459.
6. Р о и т б* а к А. И. Труды Ин-та физиол. АН ГССР, 1953, 9, 97.
7. HanberyJ. a. Jasper H. J. Neurophysiol., 1953, 16, 252.
8. Б е р и т о в И. С. Общая физиология нервной и мышечной системы, II. изд.-во «Медицина», 1966, 288.
9. Б е х т е р е в В. М. Проводящие пути спинного и головного мозга, ч. 2. СПб., 1898.
10. О •' L е а г у J. Н. а. В i sh о р G. H. J. Compar. Neurol., 1938, 68, 423.
11. П о л я к о в Г. И. Архив анат., гистол. и эмбриол., 1953, 30, 48.
12. Беритов И. С., БрегадзеА. Н. иЦкипуридзе Л. Сообщ. АН ГССР, 1942, 3, № 2.
13. Беритов И. С. иГедеваниД. М. Труды Ин-та физиол. АН ГССР, 1945, 6.
14. L о г е n t е d е N 6. R. Cerebral cortex. In: F u 11 о n J. F. «Physiology of nervous system», 1938, 274.
15. S h о 1 1 D. A. J. Anat., 1955, 89, 31.
16. Школьник-ЯрросЕ. Г. Нейроны и межнейронные связи. Зрительный анализатор. Л., изд-во «Наука». 1965.
17. CarrerasM. a. AndersonS. J. Neurophysiol., 1963, 26, 1. 48. М о u n t с a s t 1 е V. В., J. Neurophysiol., 1957, 20, 408.
19. Р а б и н о в и ч М. Я. Журн. высш. нервн. деят., 1961, II, 405; Гагрские беседы, Тбилиси, 1962, 4.
20. Школьник-ЯрросЕ. Г. Журн. высш. нервн. деят., 1958, 8, 123; 1961, 11, 680.
21. De ValoisR. L., Jones A. E. In: «The visual system. Neurophysiology and Psychophysics» (Sympos. 1960), Berlin — Gottingen — Heidelberg, 1961, 178.
22. D e V a 1 о i s R. L. J. Gen. Physiol., 1960, 43, 115.
23. М о р е л л Фр. Физиол. журн. СССР, 1962, 48, 251.
24. X а н а н а ш в и л и М. М. Физиол. журн. СССР, 1958, 44, 10.
25. Хананашвили М. М. Экспериментальное исследование центральных механизмов зрительной функции. Л., 1962.
26. S р е г г у R. W. J. Neurophysiol., 1947, 10, 275.
27. S р е г г у R. W., М i n о г N. а. М у е г s R. E. J. Compar. Physiol. Psychol., 1955, 48, 50.
28. S р е г г у R. W. Science, 1961. 133, 1749.
29. М у e r s R. E. In: «Function of Corpus callosum». Ciba Found. Study Group. N. 20. Ed. E. G. Eltlinger, 1965, 133.
30. FesherO., HalaszP. a. MeshlerF. «Kiserl. orvostad», 1964, 16, N 3, 247.
31. Е Ь п е г F. Е. а. М у е г s R. E. Anat. Record., 1962, 142, 229; J. Compar. Neurol., 1962, 118, 1.
32. 0 ' L e a ry J. H. J. Compar. Neurol., 1941, 25, 131.
33. Scheibel M. E. Anat. Record., 1955, 121, 362; М. Е. Ш а и б е л ь и А. Б.Шайбель В кн. «Ретикулярная формация мозга». Медшз, 1962, 57.
34. D е m р s е у Е. W. а. М о г i s о n R. S. Amer. J. Physiol., 1843, 138, 283.
35. М о г u z z i G. а. М a g о u n H. W. EEG and Clin. Neurophysiol., 1949, 1, 455.
36. J a s p e r H. EEG and Clin. Neurophysiol., 1949, 1, 405; 1955, 7, 99.
37. P о и т б а к А. И. Кшвський ун-т 1м. Т. Г. Шевченка, Науков! записки, 16 и 17, 167. Сб. пссвящ. Д. С. Воронцову, Кшв, 1957; Доклады на Московском коллокв. Междунар. ЭЭГ общ-ва, 6-И августа 1958.
38. П о л я к о в Г. И. Журн. высш. нервн. деят., 1956, 6, 469.
39. К a p p e r s С. U. A r i ё n s, H u Ь е r G. С. а. С г о s Ь у Е. С. The comparative anatomy of the nervous system of vertebrates, including man., N. Y., 1936.
40. E b b e с k e U. Die korticalen Erregungen. Leipzig, 1919.
41. G о 1 d s t e i n K. Handbuch d. normakn Physiologie u. Patologie, 1927, 10, 600.
42. Б л у м е н а у Д. В. Материалы I Всесоюзн. съезда по изучению поведения, 1930, 381.
43. Б е р и т о в И. С. Индивидуально приобретенная деятельность ЦНС. Тбилиси, 1932.
44. Р е n f i e 1 d W. Amer. Res. Nerv. Dis. Proc., 1952, 30, 519; The excitable cortex in conscious man. Liverpool, 1958.
45. FlechsigP Gebrin und Scele Leipzig, 1S96.
46. Поляков Г. И. иСаркисовС. А. В кн. «Цитоархптектоника коры большого мозга человека». Медгиз, 1949, 102.
47. Г е р ш у н и Г. В. Физиол. журн. СССР, 1947, 33, 393; 1949, 35, 542.
48. Б е р и т о в И. С. Труды Ин-та физиол. АН ГССР, 1945, 6, 173.
49. Б р е г а д з е А. Н. Труды Ин-та физиол. АН ГССР, 1945, 6, 157.
Дата добавления: 2017-01-13; просмотров: 778;