В кожно-мышечной воспринимающей области
особенно обширны и многочисленны двигательные участки. Здесь имеются особые участки для движения передних конечностей и особые для задних; существуют также отдельные участки для головы, шеи, туловища, хвоста. У высших позвоночных, как кошка и собака, эти участки расположены по обе стороны крестовидной борозды (рис. 68). У человека и обезьян они расположены главным образом впереди роландовой борозды, т. е. в передней центральной извилине (рис. 69. Пенфилд и Болдри, 5).
У собак и кошек на границе передней сигмоидной извилины и префронтальных долей находится ряд двигательных участков, производящих движение глаз и поворот головы в противоположную сторону, сокращение мышц лица и приподнятие губ и другие движения, происходящие во время ориентировочных и эмоциональных реакций (Бехтерев, 7).
С развитием класса млекопитающих количество и дифференциация двигательных участков растут. Дифференциация идет так далеко, что каждое движение любой конечности можно вызвать из особого участка. Так, например, можно найти такие точки, откуда по преимуществу вызывается сгибание данной ноги, а также такие, которые дают разгибание (Шеррингтон и Геринг, 7, рис. 70). Особенно многочисленны и тонко дифференцированы двигательные участки у высших обезьян и у человека. При раздражении перед-
|
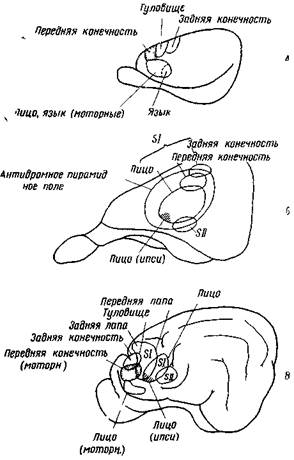
Рис. 68. Двигательные и сенсорные зоны в сенсомоторной коре больших полушарий крысы (А), кролика (Б) и кошки (В)
Сенсорная первичная (SI) и сенсорная вторичная зоны (SII) в некоторой мере перекрываются двигательной зоной, установленной антидромным раздражением пирамидного пути в продолговатом мозге. Для мозга крысы даны только двигательные участки (Терцуоло п Эди)
ней центральной извилины у обезьян было получено около 400 разных движений (Лейгон, Шеррингтон, 8). Из этой зоны могут вызываться изолированные движения не только отдельных конечностей, но и отдельных суставов и даже отдельных мышц (Ц. Фохт и О. Фохт, 9). На этом основывается возможность производства тонких рабочих движений человека, так называемых инструментальных движений.
У человека при электрическом раздражении коры можно вызвать такие тонко дифференцированные движения, которые являются частями трудового процесса, т. е. привычными движениями, образовавшимися в результате упражнения (Бергер, 10). Это указывает на то, что в двигательной области коры мозга в результате упражнения образуется множество разнообразных нервных комбинаций, которые служат для вызова трудовых движений.
Двигательные участки кожно-мышечной области не имеют постоянного анатомического положения; они располагаются у разных людей в разных местах и с течением времени меняют положение даже у одного и того же человека. Но представительство каждого двигательного органа не выходит из определенных границ. Эти границы для главных органов довольно хорошо совпадают с точками их ощущений (было показано на рис. 53, Б и В). Пенфилд и Болдри, по наблюдениям которых составлен этот рисунок, обнаружили, что чем больше пользуется человек тем или другим двигательным органом, тем чаще встречаются соответствующие двигательные точки в коре большого мозга. В этом отношении на первом месте стоит рука, вернее кисть, затем губы и локтевой сустав. В отношении чувствований также больше всега точек находят для руки, а затем для губ и языка.
Историческая справка. Двигательные участки в коре больших полушарий впервые были обнаружены Фричем и Гитцигом (1870,11,12). Они установили на собаках существование в крестовидной области коры таких участков, электрическое раздражение которых дает сокращение мышц конечностей и других частей тела. После них Феррье обнаружил, что вызванные при раздражении коры двигательные эффекты являются определенными, вполне координированными актами (1873, 13). Лучиани и Тамбурини (1878, 14) определили положение возбудимых областей специально в отношении каждого движения. Они жевпервые установили, что эти области не совсем одинаковы у разных индивидов или дажев разных полушариях одного и того же животного. Соответствующие опыты на обезьянах впервые проделал Феррье (1877), а затем Бифор и Хорслей (1887, 15); наконец, на
|
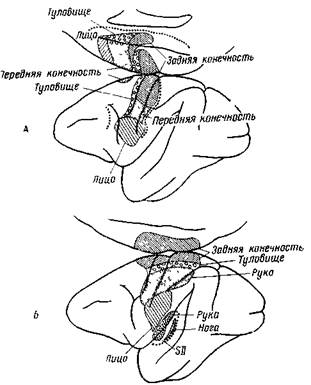
Рис. 69. Двигательные (А) и сенсорные (Б) зоны в сенсомоторной коре у обезьяны. Зоны на медиальной поверхности полушарий изображены отдельно сверху. Находящиеся в глубине борозды участки обозначены линией из кружков. Верхняя часть двигательной зоны простирается до верхнего края поясной извилины. Сенсомоторная область (S II) проходит за сильвиеву борозду (Терцуоло и Эди)
орангутанах Шеррингтон иГрюнбаум (16) установили, что у высших обезьян двигательные точки существуют только в передней центральной извилине и в глубине центральной, т. е. роландовой борозды, что они отсутствуют в задней центральной извилине. Однако на основании опытов всех поименованных исследователей известно, что двигательные участки не ограничиваются описанной областью, а встречаются и вне этих пределов в разных долях больших полушарий.
На людях соответствующие экспериментальные исследования производили Феррье, Бифор и Хорслей (1890), Бехтерев (1899, 17) и др. Наблюдения были случайными, во время различных хирургических операций на голове, но ими было установлено полное сходство большого мозга человека и высших обезьян в отношении локализации двигательных участков.
При электрическом раздражении поверхности двигательной зоны движения возникают не потому, что раздражение действует прямо на проекционные пирамиды. Последние лежат в глубине коры ниже четвертого слоя и они прямо раздражаются только при очень большой интенсивности электрического тока. Электрический ток первым долгом возбуждает нервные элементы поверхностного слоя. Эти последние приводят в активное состояние определенные нервные комплексы, состоящие из вставочных пирамидных

Рис. 70. Двигательные участки в коре головного мозга собаки
А — вид мозга сверху; Б — вид мозга сбоку. S — крестовидная борозда; F — сильвиева борозда; О — обонятельная луковица. Двигательные участки: 1 — для мышц шеи и затылка, 2 — разгибания и отведения передней ноги, з — сгибания и приведения передних ног, 4 — движения задней ноги, 5 — движения лица, в — бокового движения хвоста, г — отгибания назад передней ноги, 8— поднятия плеча и перестановки ноги вперед (шагание), S — закрытия век, а — поднятия углов рта, Ь — раскрытия рта и движения языка, d — раскрытия век, ее — platisma (по Ландуа)
и передаточных звездчатых нейронов верхних слоев коры. Они образуют нервные круги коры, которые и активируют проекционные пирамиды. Как уже указывалось в предыдущей главе, проекционные пирамиды не должны входить в эти нервные круги. Они активируются коллатералями из этих кругов непосредственно или через вставочные нейроны (рис. 65, 66). Видимо, для каждого движения существует свой нервный комплекс, свой нервный круг, который образуется в индивидуальной жизни. И если двигательные точки занимают разное положение у отдельных особей и меняют свое положение в индивидуальной жизни, то, конечно, это происходит не благодаря изменениям положения проекционных пирамидных нейронов, а благодаря образованию означенных нервных кругов из разных вставочных пирамидных и звездчатых нейронов.
Итак, в каждой воспринимающей области существуют участки, искусственное раздражение которых производит совершенно такое же движение, какое обычно вызывается при возбуждении этой воспринимающей области в ответ на раздражение соответствующего рецептора. Эти участки представляют определенные функционально связанные единицы из вставочных пирамидных и передаточных звездчатых нейронов верхних слоев коры. Они со своей стороны активируют определенные проекционные пирамидные нейроны.
Дата добавления: 2017-01-13; просмотров: 915;