Схема 29. Реакции гиперчувствительности IV типа — гиперчувствительность замедленного типа
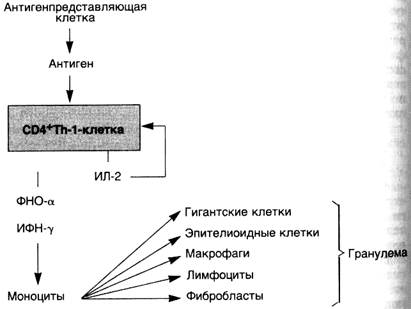
При персистенции антигена макрофаги часто трансформируются в эпителиоидные клетки, которые окружает вал из лимфоцитов — формируется гранулема. Такой тип воспаления характерен для реакций гиперчувствительности IV типа и называется гранулематозным воспалением.
Последовательность событий при ГЗТ можно рассмотреть на примере туберкулиновой реакции, которая начинается с первой встречи индивидуума с микобактериями туберкулеза. CD4+T-лимфоциты распознают белки туберкулезной микобактерий, связанные с молекулами II класса на поверхности моноцитов или других антигенпредставляющих клеток. Эта первая встреча вызывает дифференцировку СD4+Т-клеток в Th-1-клетки. Образование Th-1 -клеток является важным звеном в патогенезе ГЗТ, так как ее дальнейшее развитие зависит от цитокинов, которые секретируются Th-1-клетками. У неиммунизированного хозяина микроорганизмы впервые подвергаются атаке макрофагами, что приводит в некоторых случаях к продукции ИЛ-12. Этот цитокин регулирует дифференцировку СD4+Т-клеток в Th-1. Таким образом, развитие ГЗТ может быть связано с продукцией ИЛ-12. Сенсибилизированные Th-1-клетки поступают в кровоток и остаются там длительное время, иногда годами. В ответ на инъекцию туберкулина индивидууму, который имел раньше контакт с микобактериями туберкулеза, сенсибилизированные Th-1-клетки взаимодействуют с антигеном на поверхности антигенпредставляющих клеток и активируются, например подвергаются бласттрансформации и пролиферации. Эти изменения сопровождаются секрецией цитокинов, ответственных за ГЗТ.
ИФН-7 является наиболее важным медиатором ГЗТ и сильным активатором макрофагов. Активированные макрофаги обладают способностью к фагоцитозу и уничтожают микроорганизмы. На поверхности макрофагов имеются молекулы II класса, с помощью которых они способствуют представлению антигена. Возрастает их активность, направленная на элиминацию опухолевых клеток. Макрофаги секретируют некоторые полипептидные факторы роста, такие как тромбоцитарный фактор роста и трансформирующий фактор роста. Последний стимулирует пролиферацию фибробластов и усиливает синтез коллагена. Таким образом, активированные макрофаги обеспечивают элиминацию антигена, а если активация продолжается, то способствуют развитию фиброза и его отграничению.
ИЛ-2 вызывает аутокринную и паракринную пролиферацию Т-клеток, которые аккумулируются в местах развития ГЗТ, причем включение в инфильтрат антигенспецифических CD4+T-лимфоцитов активируется ИЛ-2.
ФНО-а и лимфотоксин являются цитокинами, оказывающими воздействие на эндотелиальные клетки. Повышенная секреция простациклина эндотелиальными клетками приводит к увеличению кровотока в результате вызываемого им расширения сосудов. Наблюдается усиление экспрессии адгезивной молекулы Е-селектина (ELAM-1), что способствует прикреплению пришлых лимфоцитов и моноцитов. Происходит усиление секреции низкомолекулярных хемотаксических факторов, например ИЛ-8. Все эти изменения в эндотелии способствуют выходу лимфоцитов и моноцитов за пределы сосудистого русла в места развития реакции ГЗТ. ФНО-а также играет важную роль в образовании гранулем.
Этот тип гиперчувствительности является главным механизмом защиты от различных внутриклеточных патогенных агентов, включая микобактерии, грибы и некоторых паразитов, а также может участвовать в отторжении трансплантата и опухолевом иммунитете.
Цитотоксичность, опосредованная Т-лимфоцитами.При данной цитотоксичности сенсибилизированные СD8+Т-лимфоциты уничтожают клетки-мишени, которые являются носителями антигена. Такие эффекторные клетки называют цитотоксическими Т-лимфоцитами (ЦТЛ). Т-лимфоциты, направленные против антигенов гистосовместимости на поверхности клеток, играют важную роль в отторжении трансплантата. Они также участвуют в защите от вирусных инфекций. В клетках, пораженных вирусом, вирусные пептиды связываются с молекулами I класса и в виде комплексов транспортируются к поверхности клетки. Этот комплекс распознается CD8+ЦТЛ. Лизис зараженных клеток завершается до репликации вируса, что приводит к уничтожению инфекции. Полагают, что многие опухолевые антигены также могут быть представлены на поверхности клеток, а ЦТЛ могут участвовать и в противоопухолевом иммунитете.
Отторжение трансплантата
В отторжении трансплантата принимают участие некоторые реакции гиперчувствительности (схема 30). Реакция отторжения зависит от распознавания хозяином пересаженной ткани как чужеродной. Антигенами, ответственными за такое отторжение у человека, являются антигены главного комплекса гистосовместимости (HLA). Отторжение трансплантата — сложный процесс, во время которого имеют значение как клеточный иммунитет, так и циркулирующие антитела.
Схема 30. Отторжение трансплантата
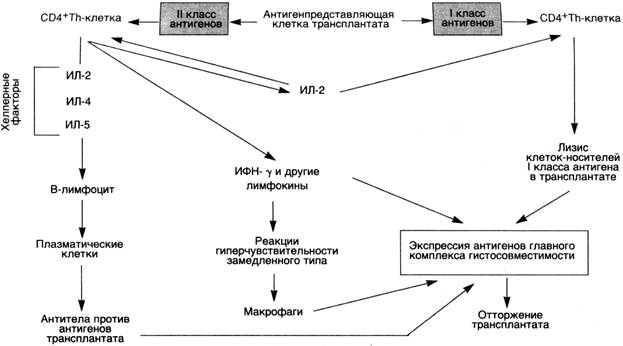
Реакции, обусловленные Т-лимфоцитами.Активированные СD4+Т-хелперы участвуют в активации CD8+ ЦТЛ. Развитие реакций, опосредованных Т-лимфоцитами, происходит при встрече лимфоцитов реципиента с антигенами HLA донора. Полагают, что наиболее важными иммуногенами являются дендритические клетки в донорских органах. Т-клетки хозяина встречаются с дендритическими клетками в пересаженном органе, а затем мигрируют в регионарные лимфатические узлы. Предшественники СШ+ЦТЛ (прекиллеры Т-клеток), обладающие рецепторами к HLA-антигенам I класса, дифференцируются в зрелые ЦТЛ. Процесс дифференцировки сложен и не совсем понятен. В нем участвуют антигенпредставляющие клетки, Т-лимфоциты и цитокины ИЛ-2, ИЛ-4 и ИЛ-5. Зрелые ЦТЛ лизируют пересаженную ткань. Кроме специфических ЦТЛ, образуются лимфокинсекретирующие СО4+Т-лимфоциты, которые играют исключительно важную роль в отторжении трансплантата. Как и при замедленной реакции гиперчувствительности, активированные СО4+Т-лимфоциты выделяют цитокины, вызывающие повышение сосудистой проницаемости и местное скопление мононуклеарных клеток (лимфоцитов и макрофагов). Считают, что ГЗТ, проявляющаяся повреждением микрососудов, ишемией и деструкцией тканей, является наиболее важным механизмом деструкции трансплантата. Более вероятно, однако, что относительная важность цитотоксичности, связанной с СБ8+Т-лимфоцитами, по сравнению с реакциями, обусловленными СО4+Т-лимфоцитами, зависит от природы несоответствия HLA донора и HLA реципиента.
Реакции, обусловленные антителами.Эти реакции могут протекать в двух вариантах. Сверхострое отторжение развивается в том случае, если в крови реципиента имеются антитела против донора. Такие антитела могут встречаться у реципиента, у которого уже было отторжение трансплантата. Предшествующие переливания крови от HLA-неидентифицированных доноров также могут привести к сенсибилизации вследствие того, что тромбоциты и лейкоциты особенно богаты HLA-антигенами. В таких случаях отторжение развивается немедленно после трансплантации, так как циркулирующие антитела образуют иммунные комплексы, которые оседают в эндотелии сосудов пересаженного органа. Затем происходит фиксация комплемента и развивается реакция Артюса.
У реципиентов, которые не были предварительно сенсибилизированы к антигенам трансплантата, экспозиция донорских HLA-антигенов I и II класса сопровождается образованием антител. Антитела, образованные реципиентами, могут вызывать повреждение с помощью нескольких механизмов, в том числе комплементзависимой цитотоксичности, антителозависимого, обусловленного клетками цитолиза и выпадения комплексов антиген — антитело. Изначально мишенью для этих антител служат сосуды трансплантата, поэтому феномен антителозависимого отторжения (например, в почке) представлен васкулитом.
Лекция 18
Дата добавления: 2016-12-26; просмотров: 1768;