Адаптивные реакции организма человека к жаркой среде обитания
У находящегося в покое человека высокая тепловая нагрузка за 30 м вызывает подъем температуры кожи до 40°С, а ректальной температуры — до 39°С. Под действием атмосферного тепла раздражаются температурные рецепторы кожи, которые вместе с повышающейся температурой циркулирующей крови активируют центральные термочувствительные структуры (в гипоталамусе). Активация последних вызывает изменение посылки симпатической адренерги-ческой импульсации к сосудистым регионам оболочки тела и внутренних органов. При этом симпатическая адренергическая активность снижается в вазоконстрикторных волокнах кожи, что вызывает раскрытие артерио-венозных анастомозов, снижение тонуса вен, а также подавляет нейрогенный тонус артериальных сосудов кожи (т.е. развивается пассивная вазодилатация). При дальнейшем повышении температуры тела включаются механизмы активной нейрогенной дилатации сосудов кожи, обусловленные активацией специализиро-
282


 ванных вазодилататорных или судомоторных нервов. Активная дила-тация сосудов на 90% увеличивает кровоток во время теплового стресса и обеспечивает более эффективную теплоотдачу в коже, чем пассивная вазодилатация. В результате пассивной и активной вазо-дилатации расширяются артериальные и венозные сосуды кожи и слизистых верхних дыхательных путей, что увеличивает теплоотдачу во внешнюю среду путем теплопроведения, излучения и увеличения снабжения потовых желез кровью. Кожный кровоток при этом увеличивается до 8 л/мин по сравнению с 0,5 л/мин при термонейтральном диапазоне (рис.24.1). Вазодилатация в пальцах и кистях рук начинается уже при температуре воздуха, равной +22°С, в стопе — при +30°С. При этом кровоток в верхних конечностях повышается в большей мере, чем в нижних. Так, в сауне у человека кровоток через кисть может возрастать в 30 раз, а через дистальные фаланги пальцев рук — в 70-100 раз по сравнению с термонейтральным диапазоном среды.
ванных вазодилататорных или судомоторных нервов. Активная дила-тация сосудов на 90% увеличивает кровоток во время теплового стресса и обеспечивает более эффективную теплоотдачу в коже, чем пассивная вазодилатация. В результате пассивной и активной вазо-дилатации расширяются артериальные и венозные сосуды кожи и слизистых верхних дыхательных путей, что увеличивает теплоотдачу во внешнюю среду путем теплопроведения, излучения и увеличения снабжения потовых желез кровью. Кожный кровоток при этом увеличивается до 8 л/мин по сравнению с 0,5 л/мин при термонейтральном диапазоне (рис.24.1). Вазодилатация в пальцах и кистях рук начинается уже при температуре воздуха, равной +22°С, в стопе — при +30°С. При этом кровоток в верхних конечностях повышается в большей мере, чем в нижних. Так, в сауне у человека кровоток через кисть может возрастать в 30 раз, а через дистальные фаланги пальцев рук — в 70-100 раз по сравнению с термонейтральным диапазоном среды.
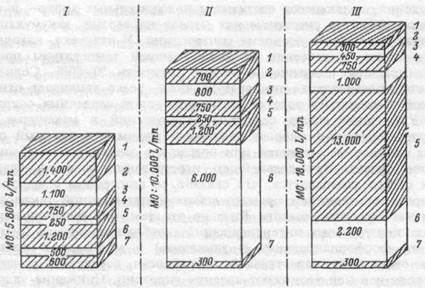
Рис.24.1.Распределение минутного объема крови в тканях.
I — в условиях покоя в термонейтрапьном диапазоне,
II — в условиях покоя, но в жаркой среде,
III — в жаркой среде, во время физической работы.
1 — спланхнический кровоток, 2 — почечный,
3 — церебральный, 4 — коронарный, 5 — в скелетных мышцах, 6 — кожный, 7 — в других тканях.
 При тепловом стрессе увеличивается посылка симпатической ад-ренергической активности от термочувствительных структур гипоталамуса к сердцу и сосудам внутренних органов, оказывая положительный хроно- и инотропный эффект, опосредуемый через
При тепловом стрессе увеличивается посылка симпатической ад-ренергической активности от термочувствительных структур гипоталамуса к сердцу и сосудам внутренних органов, оказывая положительный хроно- и инотропный эффект, опосредуемый через
283
 В2- адренорецепторы на сердечную мышцу, и вызывая сокращение а- адренореактивных артериальных и венозных сосудов чревной области (кишечника, желудка, печени и др.), а также артериальных сосудов почек. Усиление резистивной функции сосудов в этих регионах при гипертермии снижает в них кровоток и компенсирует сдвиг последнего в места теплоотдачи.
В2- адренорецепторы на сердечную мышцу, и вызывая сокращение а- адренореактивных артериальных и венозных сосудов чревной области (кишечника, желудка, печени и др.), а также артериальных сосудов почек. Усиление резистивной функции сосудов в этих регионах при гипертермии снижает в них кровоток и компенсирует сдвиг последнего в места теплоотдачи.
Рост кровотока в коже обеспечивается, в основном, увеличением сердечного выброса и, в меньшей степени, его перераспределением между "ядром" и "оболочкой" тела. Так, при тепловой нагрузке на организм человека, находящегося в состоянии физического покоя, снижение кровотока в сосудах чревной области, почках и скелетных мышцах "освобождает" до 1 л крови/мин, остальная же часть увеличивающегося кожного кровотока — до 6- 7 л/мин обеспечивается приростом сердечного выброса. Его увеличение у людей при тепловом воздействии идет параллельно повышению температуры тела и подчиняется требованиям терморегуляции и интенсивности реакций, контролирующих транспорт тепла. В основе роста величины сердечного выброса лежит, главным образом, увеличение числа сердечных сокращений, являющееся следствием положительных хроно- и ино-тропных эффектов симпатических нервов на сердце, аккумуляции в миокарде адреналина во время гипертермии. У человека, находящегося в покое, в условиях высокой внешней температуры прирост числа сердечных сокращений может достигать 50-80%. Солнечная радиация, повышающая температуру кожи, резко усиливает влияние высокой температуры воздуха на прирост числа сердечных сокращений. Увеличение числа сердечных сокращений в механизме повышения сердечного выброса является ведущим, т.к. ударный объем сердца вначале не меняется или возрастает лишь в первые минуты действия тепла. При дальнейшем перегревании человека ударный объем сердца уменьшается, что связано со снижением сократимости миокарда под влиянием прямого действия повышенной температуры крови на сердечную мышцу. Вызвано это тем, что увеличивающаяся вначале перегревания интенсификация аэробных процессов (окислительного фосфорилирования и гликолиза) в миокарде в последующем отстает от нарастающей потребности сердечной мышцы в кислороде и в ней возникают явления гипоксии. Снижение ударного объема связано не только с метаболическими нарушениями в миокарде, но и с уменьшением венозного возврата крови к сердцу, что снижает величину давления наполнения сердца.
Высокая температура воздуха, действующая на неадаптированного к жаре человека, вызывает фазные изменения артериального давления. В первую из них, уже при небольшом увеличении температуры тела, снижаются систолическое и диастолическое давление. Основной причиной снижения диастолического давления является уменьшение общего периферического сопротивления сосудов. Снижение последнего начинается при температуре воздуха, равной +28°С. Наиболее значимо общее периферическое сопротивление сосудов снижается при повышении температуры тела до +38°С, составляя 40- 55% от исходной величины. Дальнейшее повышение температуры тела (вто-
284


 рая фаза), напротив, может характеризоваться повышением общего периферического сопротивления сосудов и диастолического давления при выраженном снижении систолического давления. Повышение общего периферического сопротивления сосудов и диастолического давления является следствием компенсаторных сосудистых реакций, направленных на поддержание уровня системного артериального давления и необходимого возврата крови к сердцу. Снижение систолического давления является следствием понижения ударного объема сердца.
рая фаза), напротив, может характеризоваться повышением общего периферического сопротивления сосудов и диастолического давления при выраженном снижении систолического давления. Повышение общего периферического сопротивления сосудов и диастолического давления является следствием компенсаторных сосудистых реакций, направленных на поддержание уровня системного артериального давления и необходимого возврата крови к сердцу. Снижение систолического давления является следствием понижения ударного объема сердца.
В условиях высокой внешней температуры наиболее ранней реакцией, вызывающей перераспределение региональных объемов крови, является расширение кожных вен, влекущее функциональное депонирование значительного количества крови, которое практически выключается из общей циркуляции. Это связано с тем, что в венозных сплетениях кожи кровь движется в 5-20 раз медленнее, чем в магистральных венах. Увеличение объема венозного русла кожи компенсируется снижением у людей центрального и легочного объемов крови (на 11-22%). Однако, в результате депонирования крови в кожных венах снижается эффективный объем циркулирующей крови — возникает относительная гиповолемия. Ее компенсация достигается за счет перемещения части внеклеточной жидкости в сосудистое русло. Так, уже в первые 30 минут теплового воздействия на организм человека у него увеличивается объем плазмы крови. Вода поступает в кровь из межклеточного пространства внутренних органов — печени, легких и др. Повышенный объем плазмы при гипертермии может сохраняться долго, если человек не ограничивает потребление воды. Важную роль в этом процессе играет увеличение коллоидно-осмотического давления плазмы крови, задерживающего воду в сосудистом русле, а также уменьшение выведения воды с мочой.
Симпатическое возбуждение, возникающее при действии высокой температуры на организм, вызывает сужение почечных сосудов и, как следствие, уменьшение почечного кровотока и скорости клубочковой фильтрации. В результате, выделительная функция почек снижается. Так, диурез уменьшается в жару до 300-600 мл в сутки (при норме около 1,5 литров). Однако, на снижение выделительной функции почек влияет и повышенная секреция гормонов — вазопрессина и альдос-терона. Увеличение секреции первого связано: 1) с рефлекторным ответом нейрогипофиза на снижение центрального венозного давления; 2) с раздражением осморецепторов, вызванным увеличением осмотической концентрации крови потерями воды с потом. Возросшая секреция вазопрессина усиливает реабсорбцию воды в почках, что повышает объем циркулирующей крови. Симпатическое возбуждение при гипертермии снижает кровоток в почках, активирует ренин-ан-гиотензин-альдостероновую систему. Так, у человека 20 минут пребывания в сауне увеличивает в плазме крови содержание ренина на 95%, ангиотензина — на 195% и алъдостерона — в 5 раз. Активация ренин- ангиотензин-альдостероновой системы, в свою очередь, усиливает почечную и чревную вазоконстрикции, дополняя эффект симпати-
285

 ческой активации, адресованной этим сосудистым зонам. Секрецию альдостерона стимулирует и уменьшение хлористого натрия в крови, вызванное потерями последнего с потом.
ческой активации, адресованной этим сосудистым зонам. Секрецию альдостерона стимулирует и уменьшение хлористого натрия в крови, вызванное потерями последнего с потом.
Увеличение кровотока в коже обеспечивает потовые железы кровью в количестве, необходимом для повышенного образования ими секрета. У неадаптированного к жаре человека, находящегося в состоянии физического покоя, воздействие внешнего тепла, повышая температуру "ядра" тела до 38°С, повышает секрецию пота до 10-12 мл/мин (при комнатной температуре она составляет 600 мл/ день). У адаптированного к жаре человека, также находящегося в состоянии физического покоя, потоотделение достигает 20 мл/мин. Это связано с более быстрым усилением кровотока в коже и с повысившейся способностью к потоотделению потовых желез у адаптированных к жаре людей. Неакклиматизированные к жаре люди в первые дни действия тепла на организм теряют 15-30 г солей в день с потом и мочой, но после 4-6 недель их акклиматизации потери солей уменьшаются до 3-5 г в день. Лучшее сохранение хлористого натрия в организме связано с повышенной секрецией альдостерона.
При высокой температуре окружающей среды дыхание учащается, легочная вентиляция увеличивается, в основном, за счет глубины дыхания. Значительная гипервентиляция легких, способствуя вымыванию угольной кислоты из крови, понижает в ней напряжение СО2, что, в свою очередь, смещает рН крови в щелочную сторону. Поэтому гипокапния и дыхательный алкалоз являются постоянными признаками перегревания организма человека. Способность крови связывать СО2 снижается и под влиянием роста ее температуры.
Уменьшение кровоснабжения желудочно-кишечного тракта, вызванное перераспределением крови к кожным сосудам при повышении температуры тела, ведет к снижению секреторной, всасывательной и моторной функций органов пищеварения, понижению аппетита. Так, при повышении температуры тела уменьшается секреция слюны, в ней повышается содержание плотного остатка, что связано с потерей воды организмом при испарительном термолизисе. При потере воды, равной 8% от массы тела, слюноотделение прекращается полностью. Гипертермия снижает объем секреции желудочного сока и содержание в нем соляной кислоты и пепсина. Это, в свою очередь, уменьшает переваривающую силу желудочно-кишечного сока. Повышение температуры тела у человека до 37,5°С угнетает моторику желудка. В результате, замедляется эвакуация из него пищи. Под влиянием высокой внешней температуры на организм у неадаптированных людей уменьшается также объем секретируемого панкреатического сока, в нем снижается содержание протеолитичес-ких ферментов. Эвакуация желчи из желчного пузыря уменьшается вследствие снижения тонуса его стенок. Это приводит к застою желчи. По-видимому, вследствие указанных сдвигов у людей при тепловых воздействиях ухудшается аппетит, исчезает желание принимать пищу животного происхождения. Существующий у жителей жарких стран обычай выпивать перед едой некоторое количество


 жидкости (чая и др.), сопутствующий приему пищи отдых, уменьшение тепловой нагрузки, по-видимому, не только уменьшают дегидратацию организма, но и способствуют перемещению части крови к желудочно-кишечному тракту.
жидкости (чая и др.), сопутствующий приему пищи отдых, уменьшение тепловой нагрузки, по-видимому, не только уменьшают дегидратацию организма, но и способствуют перемещению части крови к желудочно-кишечному тракту.
Тепловой стресс, активируя симпатоадреналовую и гипофизарно-адренокортикальную системы, вызывает усиление секреции катехо-ламинов мозговым веществом надпочечников и глюкокортикоидов их корой. Под влиянием этих гормонов в печени усиливается образование глюкозы в ходе гликолиза, стимулируется глюконеогенез.
У акклиматизированных к жаре людей реакция нейроэндокринной системы на тепловые воздействия снижается. В результате, уменьшается и концентрация в плазме катехоламинов, глюкокортикоидов, тироксина. Поэтому у адаптированных к жаре людей замедляется глюконеогенез, снижается скорость гликолиза, восстанавливаются запасы гликогена в печени и скелетных мышцах. В целом, перегревание организма приводит к понижению энергетического обмена.
Снижение кровотока во внутренних органах в условиях нарастающей гипертермии приводит к снижению отношения доставки кислорода к его потреблению в тонком кишечнике, печени, почках, скелетных мышцах. В результате, в этих органах возникает тканевая гипоксия, что активирует образование энергии в ходе анаэробного гликолиза (глава 1), и приводит к увеличению концентрации недо-окисленных продуктов обмена (молочной кислоты и др.) в этих тканях и крови.
Система крови отвечает на перегревание организма значительными изменениями. Тучные клетки и базофильные лейкоциты реагируют на повышение температуры крови быстрой дегрануляцией, секретируя в кровь гепарин, гистамин и другие биологически активные вещества. При повышении температуры "ядра" тела уровень гистамина в крови увеличивается в несколько раз. У адаптированных к жаре людей активируются и механизмы, препятствующие росту гистамина в крови, нарастает содержание гистопексина и активность гистаминазы, понижающих концентрацию в ней гистамина. Это снижает проницаемость кровеносных сосудов, повышаемую гистами-ном, снижает его вазодилататорный эффект на артериолы брюшной полости и других регионов.
Под действием тепла в клетках тканей внутренних органов, скелетных мышцах повышается проницаемость мембран лизосом, что вызывает выход лизосомальных протеиназ в кровь. Однако, с адаптацией организма к жаре в плазме крови повышается уровень а1-антитрипсина — мощного ингибитора протеолитических ферментов, что предохраняет белки и клеточные элементы крови от избыточного протеолиза.
У людей, выходцев из умеренной зоны Земли, но живущих в жарких влажных тропиках, гематологические сдвиги (лейкопения, относительная и абсолютная нейтропения, лимфоцитоз) в целом сходны с изменениями крови у аборигенов тропиков. У европейцев, адаптированных к тропикам, и у рабочих, многие годы подвергавшихся тепловым воздействиям, возникают изменения в эритроне:
287
 нарастает гемоглобинизация и гидратация отдельного эритроцита, увеличивается содержание гемоглобина в крови, эритроцитарный уровень удерживается на значениях, приближающихся к нижней границе европейской нормы. Уровень эритропоэтина в крови снижается и это объясняет, почему у адаптированных к жаркому климату людей происходит замедление регенерации красной крови, например, после кровопотери.
нарастает гемоглобинизация и гидратация отдельного эритроцита, увеличивается содержание гемоглобина в крови, эритроцитарный уровень удерживается на значениях, приближающихся к нижней границе европейской нормы. Уровень эритропоэтина в крови снижается и это объясняет, почему у адаптированных к жаркому климату людей происходит замедление регенерации красной крови, например, после кровопотери.
Особенности адаптации человека к работе в жаркой среде.Потребление кислорода и температура "ядра" тела закономерно нарастают во время работы возрастающей мощности. Так, работа мощностью до 50% от величины максимального потребления кислорода (МПК), производимая в комфортной температурной среде, увеличивает температуру ядра до 37,4°С, а работа с 75% потреблением кислорода от величины МПК, повышает ее до 38,5°С. Температура кожи при этом остается относительно неизменной, что обеспечивает эффективную отдачу тепла. Однако, при наличии внешней тепловой нагрузки, тепло, образующееся в работающих мышцах, может еще более увеличивать температуру тела, если нарушится эффективность охлаждающего испарительного механизма и обеспечивающих его сосудистых компенсаторных реакций. Работа человека на жаре требует от системы кровообращения выполнения двух задач: 1) обеспечить мышцы кислородом, чтобы поддержать в них возросший расход энергии; 2) обеспечить перенос кровью метаболического тепла от глубоких тканей к оболочке тела. При этом кровоток в мышцах повышается до 13 л/мин, кожный — до 2,2 л/мин, а сердечный выброс — до 18 л/мин (рис.24.1), что обеспечивается за счет уменьшения кровотока в висцеральных органах. Например, в желудочно-кишечном тракте и почках кровоток снижается в этом случае соответственно до 300 и 450 мл/мин. Вазоконстрикция в висцеральных органах повышает общее периферическое сопротивление сосудов, что поддерживает на оптимальном уровне артериальное давление крови во время работы на жаре, обеспечивает достаточную перфузию сосудов головного мозга, препятствует развитию церебральной ишемии и артериальной гипотензии. Коронарный кровоток при работе на жаре может возрастать в четыре раза по сравнению с покоем.
Физическая нагрузка, даже хорошо переносимая в жару, выполняется с нарастанием доли анаэробного метаболизма во внутренних органах, что приводит к быстрому уменьшению запасов гликогена в этих тканях, нарастанию в них молочной кислоты. Этому способствует сниженная экстракция кислорода из крови в мышцах, т.к. несмотря на возрастающий в них кровоток, большое количество крови из мышц шунтируется, не отдав кислород, к сосудам оболочки тела для рассеивания тепла. Росту уровня молочной кислоты в крови способствует и сниженное поступление крови в печень из- за значительного уменьшения печеночного кровотока. Накапливающиеся в крови кислые метаболиты истощают буферные основания, создавая условия для развития метаболического ацидоза.
288
 Потери воды при работе в жару, например, при очень жаркой погоде в пустыне, могут достигать 8- 12 л/сутки с потом, при этом интенсивное потоотделение в течение нескольких часов может привести к "усталости" потовых желез, вызванной истощением их секреторной функции. Это резко нарушает способность регулировать температуру "ядра" тела. Потери жидкости при потоотделении не столь эффективно охлаждают кожу во время работы в жару при высокой влажности, когда испарительное охлаждение кожи снижено высоким давлением водяных паров в окружающем воздухе. Поэтому интенсивное потоотделение в этих условиях, хотя и приводит к большим потерям жидкости, но сопровождается минимальным испарением и охлаждением поверхности тела, повышая риск перегревания организма.
Потери воды при работе в жару, например, при очень жаркой погоде в пустыне, могут достигать 8- 12 л/сутки с потом, при этом интенсивное потоотделение в течение нескольких часов может привести к "усталости" потовых желез, вызванной истощением их секреторной функции. Это резко нарушает способность регулировать температуру "ядра" тела. Потери жидкости при потоотделении не столь эффективно охлаждают кожу во время работы в жару при высокой влажности, когда испарительное охлаждение кожи снижено высоким давлением водяных паров в окружающем воздухе. Поэтому интенсивное потоотделение в этих условиях, хотя и приводит к большим потерям жидкости, но сопровождается минимальным испарением и охлаждением поверхности тела, повышая риск перегревания организма.
Значительная потеря жидкости с потом уменьшает объем плазмы и циркулирующей крови, что снижает величины конечного диасто-лического объема желудочков и систолического объема сердца, уменьшает кровоток в коже и, следовательно, потоотделение. Минутный объем не уменьшается вследствие компенсаторно нарастающего числа сердечных сокращений. В результате снижения эффективности механизмов, обеспечивающих выведение тепла из организма, увеличивается температура "ядра" тела. Следовательно, дегидратация организма уменьшает возможность систем кровообращения и терморегуляции компенсировать тепловой стресс при физической нагрузке. Поэтому поддержание на оптимальном уровне объемов плазмы и экстрацеллюлярной жидкости у человека в условиях работы на жаре сохраняет функции кровообращения и продукцию пота. А это, в свою очередь, достигается сбалансированием потери и восполнения воды. У человека, получающего достаточное количество жидкости, работоспособность в жару сохраняется на более высоком уровне, чем у работающего в состоянии дегидратации. Употребление дополнительно 400-600 мл воды за 10-20 минут до мышечной работы на жаре, задерживает развитие дегидратации, увеличивает потоотделение во время работы и препятствует значительному повышению температуры "ядра" тела.
Для рабочего, выполняющего работу средней тяжести на открытом воздухе в условиях Средней Азии, "норма" выпиваемой воды за смену составляет 4-4,5 литра. Большинство людей устойчивы к потерям 3-4% воды (от массы тела). При потере же ее 5-8% возникает чувство усталости в положении "сидя", сонливость, апатия. Потеря свыше 10% воды вызывает у человека физические и психические нарушения. При тяжелой дегидратации необходимы срочные врачебные меры: вводят внутривенно 5% раствор глюкозы, изотонический раствор хлорида натрия (до 1 л/час в начале лечения), но не более 4 литров в сутки.
Потери пота при работе на жаре могут истощать солевой запас организма на 13-15 г/сутки (2,3-3,4 г/л пота), у неакклиматизиро-ванных людей потери солей могут быть больше. Это значительно превышает их величину в ежедневном рационе, поэтому необходима добавка в пищу соли (1/3 чайной ложки поваренной соли на литр
289
 воды). Потери калия, кальция и магния при выделении пота невелики и могут быть восполнены богатой ими диетой. Например, стакан томатного сока возмещает почти весь калий, кальций и магний, содержащиеся в 2- 3 литрах пота. При значительных потерях натрия внутривенно вводят изотонический раствор хлорида натрия. При интенсивных потерях жидкости, связанных с жарой, возникает гиповитаминоз водорастворимых витаминов С и В,.
воды). Потери калия, кальция и магния при выделении пота невелики и могут быть восполнены богатой ими диетой. Например, стакан томатного сока возмещает почти весь калий, кальций и магний, содержащиеся в 2- 3 литрах пота. При значительных потерях натрия внутривенно вводят изотонический раствор хлорида натрия. При интенсивных потерях жидкости, связанных с жарой, возникает гиповитаминоз водорастворимых витаминов С и В,.
Повышение устойчивости организма к действию тепла называется тепловой акклиматизацией. Обычно акклиматизация организма заканчивается к концу второй недели теплового воздействия. Для этого требуется от 2 до 4-х часов ежедневных тепловых экспозиций. Однако, первые тепловые воздействия должны быть тренирующими, продолжительностью около 15-20 минут. Их продолжительность и интенсивность должны возрастать постепенно. Акклиматизация достигается легче, если человек во время работы на жаре имеет возможность полностью покрывать потребность в воде. Адаптированные к жаре люди легче переносят ее воздействие, сохраняют большую работоспособность, другие виды социальной активности.
24.3. Морфо-функциональные особенности коренных жителей тропиков
Большую устойчивость коренного населения тропиков к действию на организм влажного и сухого тепла объясняют морфофункци-ональными особенностями, приобретенными популяциями в ходе их длительного существования в зоне тропиков, выразившимися в формировании "тропического" адаптивного типа человека. Под адаптивным типом понимают норму биологической реакции на комплекс условий окружающей среды, обеспечивающей состояние равновесия популяции с этой средой, но имеющей внешнее выражение в морфо- функциональных особенностях популяции.
По сравнению с жителями умеренного пояса население тропических широт характеризуется пониженной плотностью тела, уменьшенным весом, увеличением длины конечностей и туловища, малой выраженностью жировой складки, большим отношением поверхности тела к его весу, что увеличивает испаряющую поверхность тела, способствует лучшему выведению тепла из организма. Большие ширина носа, ротовой щели и поверхности губ, увеличивая площадь слизистой, повышают также отдачу влаги и охлаждают вдыхаемый воздух. Узкая, высокая и длинная голова, типичная для негроидов, нагревается медленнее, чем плоская и широкая. Курчавые волосы негров содержат большое количество воздуха, что резко уменьшает теплопроводность волосяной шапки, препятствуя нагреванию кожи головы.
У жителей влажных тропиков обнаружена большая, чем у европейцев, разница между температурой "ядра" и "оболочки" тела, кистей и стоп, что способствует лучшему выведению тепла из организма. У негроидов и монголоидов, живущих в тропиках, механизмы выведения тепла с потом совершеннее, чем у европейцев: у жителей
290
тропиков число активно функционирующих потовых желез на см2 поверхности тела выше, и за равный промежуток времени они теряют больше пота, чем европейцы. Например, у жителей пустыни, австралийских аборигенов количество пота, испаряемое с поверхности тела, составляет 13,5 мл/кг веса/час против 6,6 мл/кг веса/час у европейцев, находящихся в идентичных условиях. У аборигенов влажных тропиков в ответ на температуру, превышающую 27°С, потоотделение начинается быстрее и интенсивнее, чем у европейцев.
Водный обмен у жителей тропиков адаптирован к большим объемам испаряемой жидкости. Так, у австралийских аборигенов водный обмен выше, чем у европейцев, живущих в тех же районах пустынь (95 мл/кг/сутки против 54 мл/кг/сутки) и он еще более интенсивен у аборигенов влажных жарких тропиков по сравнению с живущими здесь европейцами (119 мл/кг/сутки против 80 мл/кг/ сутки). У жителей тропиков объем внеклеточной жидкости и объем плазмы выше, чем у европейцев. Увеличенный объем внеклеточной жидкости выполняет функцию буфера, предохраняющего организм от дегидратации при потении. Коренные жители пустынных и са-ванных районов тропиков, будучи не полностью акклиматизированными к влажной жаре, приспосабливаются к ней быстрее и лучше, чем европейцы. У них выявляется меньший прирост пульса и потери жидкости с потом при пробах с нагрузкой во влажной жаркой среде, чем у европейцев. Одной из причин этого служит, по-видимому, лучшая способность коренных жителей тропиков к равномерному потоотделению на единицу площади поверхности тела. При этом потери натрия с потом у аборигенов тропиков много ниже, чем у европейцев, что связано с особенностями гормональной регуляции их электролитного и водного обмена. Так, уровень ренина плазмы у жителей пустынь и влажных тропиков в 2- 3 раза выше, чем у европейцев. Это уменьшает кровоток в почках и скорость клубочковой фильтрации. У аборигенов тропиков повышена продукция вазопрессина, что усиливает реабсорбцию воды в почках. У жителей тропиков во время работы на жаре снижена чувствительность структур центральной нервной системы к действию теплового стресса. Видимо, вследствие этого у них, по сравнению с европейцами, отсутствуют признаки стресс-реакции на жару: снижен ответ надпочечников на АКТГ, уменьшена его продукция, а также уровень кортизона в плазме. В условиях перегревания коренные жители тропиков отличаются большей устойчивостью физиологических процессов, психических и локомоторных функций.
Основной обмен у аборигенов тропиков характеризуется меньшими значениями, чем у жителей умеренного климата. Снижение основного обмена тем больше, чем выше величина среднегодовой температуры экологической среды обитания. Так, у лиц, живущих на экваторе, основной обмен приблизительно на 20% ниже, чем у обитателей субарктических районов. Причинами его снижения являются высокая температура окружающей среды, угнетающая окислительные процессы в тканях внутренних органов, низкая калорийность рациона и слабое развитие мышечной массы. Согласно рас-
291


 четам экспертов ВОЗ, человек в тропиках должен получать в среднем в день 2225 ккал ±10%. Между тем, в Верхней Вольте житель страны потребляет в среднем 1710 ккал в день. У взрослых 40-летних мужчин этой страны вес тела в среднем оказался на 10 кг меньше, чем у выходцев из нее, но живущих в Северной Америке. Меньшей была у них и окружность плеча. Для рациона большинства африканцев характерно низкое среднесуточное содержание калорий, белков, жиров и высокое — углеводов, а для жителей тропической Африки, кроме того, характерно сниженное содержание никотиновой кислоты и рибофлавина, дефицит железа, тесно связанных со многими процессами обмена. Затруднение в восполнении возникающего дефицита железа у жителей влажных тропиков связано с тем, что многие продукты, употребляемые в этом регионе (кокосовое молоке и др.), хотя и содержат большое количество железа, но в них велико и наличие соединений растительного происхождения, ингибирующих его абсорбцию.
четам экспертов ВОЗ, человек в тропиках должен получать в среднем в день 2225 ккал ±10%. Между тем, в Верхней Вольте житель страны потребляет в среднем 1710 ккал в день. У взрослых 40-летних мужчин этой страны вес тела в среднем оказался на 10 кг меньше, чем у выходцев из нее, но живущих в Северной Америке. Меньшей была у них и окружность плеча. Для рациона большинства африканцев характерно низкое среднесуточное содержание калорий, белков, жиров и высокое — углеводов, а для жителей тропической Африки, кроме того, характерно сниженное содержание никотиновой кислоты и рибофлавина, дефицит железа, тесно связанных со многими процессами обмена. Затруднение в восполнении возникающего дефицита железа у жителей влажных тропиков связано с тем, что многие продукты, употребляемые в этом регионе (кокосовое молоке и др.), хотя и содержат большое количество железа, но в них велико и наличие соединений растительного происхождения, ингибирующих его абсорбцию.
Низкое содержание в пище животного белка, а следовательно, и содержащих сульфгидрильные группы аминокислот — метионина и цистеина, способствующих всасыванию железа, а также содержание в ряде продуктов железа в неорганической форме — затрудняет его всасывание, способствуя тем самым развитию железодефицита у коренных жителей тропиков. Снижение интенсивности обменных процессов приводит к уменьшению поглощения кислорода тканями. Соответственно уменьшению основного обмена у жителей тропиков понижены и показатели функции щитовидной железы. Однако, ведущую роль в понижении ее функции играют отмеченные выше особенности пищевого рациона жителей тропиков, пониженная у определенных групп населения физическая активность, сниженная абсолютная мышечная масса тела ( в сравнении с европейцами).
Для коренных жителей тропиков характерна наследственно закрепленная избирательная способность метаболизма к диете с избытком углеводов и дефицитом белка. У сенегальцев, нигерийцев, банту, индийцев выявлено более низкое содержание триглицеридов, фосфолипидов и холестерина в крови по сравнению с жителями США и Европы, а также представителями белой расы, живущими в тропиках. Эта особенность сочетается у них с низким уровнем липопротеинов низкой плотности и высоким уровнем активности липопротеинов высокой плотности, что препятствует развитию атеросклероза коронарных артерий (см.главу 20). При этом высокий уровень липопротеинов высокой плотности у африканцев не имеет столь выраженной тенденции к уменьшению с возрастом, как у европейцев. При объяснении данного феномена важное значение придается низкой калорийности рациона жителей тропиков. Вместе с тем, наблюдение массаев, жителей Кении и Танзании, имеющих низкую частоту ишемической болезни сердца и низкий уровень общего холестерина в сыворотке крови при традиционном обильном употреблении мяса, молока и крови зебу, свидетельствует о существовании наследственного механизма угнетения синтеза эндогенного холестерина при поступлении в организм большого количества
292
экзогенных жиров. С этим связывают, в частности, намного меньшую, чем у европейцев, частоту смерти от коронарной болезни сердца у коренных жителей Африки, живущих южнее Сахары и в тропиках Южной Америки. В коронарных сосудах стариков-африканцев обнаруживают меньше атеросклеротических бляшек, их изъязвлений и кальцификаций, чем даже у молодых европейцев.
Параметры гемостаза у коренных жителей тропиков имеют отчетливые различия от величин, характерных для жителей умеренных широт. Так, у взрослых жителей тропиков содержание тромбоцитов в крови много ниже, чем у европейцев, в том числе живущих в тропиках. Например, содержание тромбоцитов в 1 мкл крови у аборигенов Сенегала и Нигерии достигает 100-220 тыс., а у живущих в этих же странах европейцев — 200-320 тыс. Причинами тромбоцитопении могут быть: 1) носительство малярийных плазмодиев; 2) спленомегалия (увеличение селезенки), вызванная хронической малярией, увеличивает внутриселезеночный пул тромбоцитов (глава 6) и оказывает тормозящее воздействие на тромбоцитопоэз в костном мозге. У жителей тропиков, в отличие от жителей умеренного пояса, более активен фактор VIII системы свертывания крови. Несмотря на высокую активность фактора VIII, время первой стадии свертывания крови у африканцев длиннее, чем у европейцев. Это связано не только с низким уровнем тромбоцитов, но и с угнетением их агрегации, высокой концентрацией в крови иммуноглобулинов различных классов. Активность фибринолитической системы у африканцев (кенийцев, банту, сенегальцев, нигерийцев) выше, чем у местного европейского населения. У аборигенов тропиков укорочено время фибринолиза, уровень плазминогена и активатора плазминогена у них выше, чем у европейцев. Эти данные объясняют весьма малую частоту эмболии легочной артерии и тромбоза глубоких вен в тропических странах по сравнению с Европой и США.
Сравнение показателей красной крови здоровых коренных жителей тропических стран и жителей Европы, Сибири и США указывает на уменьшение количества эритроцитов в 1 мкл крови, более низкое содержание ретикулоцитов и эритропоэтина в крови у жителей тропиков. Причиной этих изменений является снижение основного обмена, понижающего кислородный запрос тканей, уменьшение активности коры надпочечников, щитовидной железы, приспособление водного обмена (за счет увеличения внеклеточного и плазменного его объемов) к повышенным нуждам физической терморегуляции. К этим влияниям прибавляется низкое содержание белков в пищевом рационе и широкое распространение у жителей тропиков как скрытого, так и явного дефицита железа.
Тропические популяции — это популяции на протяжении многих поколений непрерывно подвергающиеся не только температурным, но и антигенным и инфекционным воздействиям. Поэтому организм коренных жителей тропиков оказывается высоко резистентным не только к тепловым нагрузкам, но обнаруживает повышенную сопротивляемость к возбудителям инфекционных заболеваний, в том чис-
293

 ле к одному из самых грозных инфекционных агентов в тропиках
ле к одному из самых грозных инфекционных агентов в тропиках
— к возбудителю малярии.
Для здоровых коренных жителей тропиков характерен относительно низкий уровень альбуминов и повышенное, по сравнению с европейцами, содержание в крови иммуноглобулинов G, М, Е, обеспечивающее высокую активность гуморального звена иммунитета в защите против паразитарной инфекции. Так, содержание иммуноглобулинов в сыворотке крови взрослых жителей умеренного и тропического пояса Земли (в МЕ/мл) составляет соответственно: Ig G
- 65-208, и 146-567; Ig M — 47-380 и 34-1413; Ig E - 21-67 и
368-4344.
Причину гипоальбуминемии связывают с белковой недостаточностью питания в сочетании с наследственно закрепленным относительно низким уровнем плазменных альбуминов у жителей тропиков. Высокая концентрация гамма-альбуминов у африканцев, индейцев Америки, живущих в тропическом поясе, коренного населения Индии объясняется генетическими особенностями. Эту тенденцию усугубляет постоянный антигенный прессинг, связанный с инфицированием возбудителями малярии, гельминтозов и других инфекционных и инвазивных заболеваний обитателей этих районов.
У здоровых аборигенов жарких стран, в отличие от жителей средних широт, имеет место резко выраженная лейко- и нейтропения, сочетанная с эозинофилией, относительным и абсолютным лимфо-цитозом (табл.24.1). Причина лейкопении не установлена. В отношении нейтропении предполагают наследственный ее характер. Вместе с тем, у африканцев при остром воспалении, бактериальной инфекции в крови быстро повышается содержание зрелых форм нейтрофилъных лейкоцитов, благодаря наличию у жителей тропиков более емкого костно-мозгового пула депонированных нейтрофилов, чем у европейцев.
Таблица 24.1Показатели белой крови у жителей влажных тропиков и умеренного пояса
| Лейкоциты Эозинофилы Палочкоядерные Сегментоядерные Лимфоциты Моноциты | Жители Гвинеи (Конакри) | Жители России | ||
| в 1 мм3 | в % | в 1 мм3 | в% | |
| 5460±550 4000 - 10000 (3200 - 8300) 576±109,8 11,5*1,99 100 - 250 1-4 (102 - 2440) 27+11,2 0,55±0,24 180-400 2-5 (0 - 180) 2021+251 38,6±2,59 3065 - 5600 55 - 68 (663 - 4317) 2237±140 44,1±1,8 1200 - 2800 25 - 30 (1472 - 4731) 170±66 6,35±0,4 200 - 600 6-8 (122 - 660) | ||||
294

 Увеличению числа лимфоцитов в крови, усилению пролиферации лимфоидной ткани способствует постоянная умеренная гипертермия. Повышенная пролиферация лимфоцитов поддерживается также хроническим воздействием малярийных антигенов, снижающих функцию Т-клеток супрессоров. Учитывая важную роль лимфоцитов в поддержании тканевого гомеостаза, адаптивных реакций организма и в формировании иммунного ответа допускают, что увеличение их числа в крови при действии экстремального термического фактора носит приспособительный характер.
Увеличению числа лимфоцитов в крови, усилению пролиферации лимфоидной ткани способствует постоянная умеренная гипертермия. Повышенная пролиферация лимфоцитов поддерживается также хроническим воздействием малярийных антигенов, снижающих функцию Т-клеток супрессоров. Учитывая важную роль лимфоцитов в поддержании тканевого гомеостаза, адаптивных реакций организма и в формировании иммунного ответа допускают, что увеличение их числа в крови при действии экстремального термического фактора носит приспособительный характер.
Дата добавления: 2016-12-16; просмотров: 1835;