Вазопрессин: биосинтез, транспорт и метаболизм
Водный обмен и несахарный диабет
ФИЗИОЛОГИЯ ВОДНОГО ОБМЕНА
Осмолярность плазмы и ее основной определяющий фактор, концентрация натрия в плазме, в норме поддерживаются в достаточно узких пределах. Такая стабильность достигается главным образом за счет регуляции общего содержания воды в организме и поддержания равновесия с концентрацией натрия в сыворотке. Водный баланс контролируется комплексной системой, регулирующей потребление воды с помощью центра жажды и объем мочи посредством концентрации вазопрессина в плазме (рис. 1-1). В физиологической регуляции синтеза и секреции вазопрессина участвует два параметра: осмолярность внеклеточной жидкости и давление и объем крови. Вазопрессин – основной гормон, участвующий в регуляции водного гомеостаза и осмолярности, а ренин-ангиотензин-альдостероновая система (РААС) главным образом ответственна за регулировку артериального давления и объема крови (Robinson and Verbalis, 2011). Осморегуляция секреции вазопрессина – относительно простой процесс: небольшое повышение осмолярности вызывает усиление секреции вазопрессина, а незначительное снижение осмолярности – снижение.
Нейрогипофиз
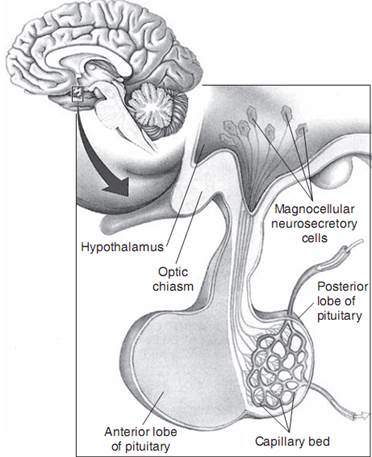
Нейрогипофиз состоит из ряда ядер гипоталамуса (супраоптических и паравентрикулярных), содержащих крупноклеточные нейроны, ответственные за синтез окситоцина и вазопрессина; аксонов этих клеток, формирующих супраоптико-гипофизарный путь; и окончаний этих клеток в задней доле гипофиза (рис. 1-2; Reeves et al, 1998). Крупноклеточные нейроны паравентрикулярных и супраоптических ядер секретируют вазопрессин или окситоцин в ответ на соответствующие стимулы. Крупноклеточные нейроны, секретирующие вазопрессин, получают нервные импульсы от различных чувствительных элементов, в том числе артериальных барорецепторов высокого давления, расположенных в сонном синусе и дуге аорты, и рецепторов объема, реагирующих на низкое давление и расположенных в предсердиях и системе легочных вен (Thrasher, 1994). Барорецепторы и рецепторы объема в норме тормозят крупноклеточные нейроны, а ослабление такого тонического торможения приводит к высвобождению вазопрессина. Сужение вен и артерий, индуцированное действием вазопрессина на рецепторы Vla кровеносных сосудов, при существующем объеме плазмы эффективно «увеличивает» объем плазмы, и секреция вазопрессина снова начинает подавляться (Robinson and Verbalis, 2011). Действие вазопрессина на почки, направленное на удержание воды, помогает восполнить объем, тем не менее, основной системой гормональной регуляции объема крови является РААС, стимулирующая реабсорбцию натрия в почках.
Вазопрессин: биосинтез, транспорт и метаболизм
Вазопрессин и окситоцин – нонапептиды, состоящие из шестичленного дисульфидного кольца и трехчленного хвоста с амидированной концевой карбоксильной группой (рис. 1-3). Аргинин-вазопрессин (АВП) – антидиуретический гормон у всех млекопитающих, кроме свиней и других членов подотряда Suina, у которых синтезируется лизиновый вазопрессин (Reeves et al, 1998). У большинства млекопитающих вазопрессин отличается от окситоцина только заменой изолейцина фенилаланином в кольце и лейцина аргинином в хвосте. Соотношение антидиуретического и сосудосуживающего эффектов вазопрессина заметно повышается при замене L-аргинина на D-
| Крупные нейросекреторные клетки
| |
| Перекрест зрительных нервов
| |
РИС. 1-2.Вид гипоталамуса и надпочечника в средне-сагиттальной проекции; видны тела крупноклеточных нейронов в гипоталамусе и их аксоны, идущие в гипофиз, где секретируются вазопрессин и окситоцин непосредственно в капилляры задней доли гипофиза. (Источник: Bear MF, et al.: Neumscience: exploring the brain, ed 3, Baltimore, 2007, Lippincott Williams & Wilkins, р. 486.)
аргинин в положении 8. В результате такой замены, а также удаления концевой аминогруппы цистеина, получается десмопрессина ацетат (ДА), синтетический препарат, имеющийся в продаже (см. рис. 1-3). ДА – препарат, применяющийся в клинической практике и обладающий более длительным и сильным антидиуретическим действием, однако не требующий введения в инъекционной форме и часто применяющийся для лечения центрального несахарного диабета (ЦНСД) у собак и кошек.
| Положение
| Окситоцин
|
| Вазопрессин
|
| ДА
|
|
|
| NH2
|
| NH2
|
|
|
|
| 1
| Цистеин
| ¾
| Цистеин
| ¾
| Цистеин
| ¾
|
| 2
| Тирозин
| S
| Тирозин
| S
| Тирозин
| S
|
| 3
| Изолейцин
|
| Фенилаланин
|
| Фенилаланин
|
|
| 4
| Глютамин
|
| Глютамин
|
| Глютамин
|
|
| 5
| Аспарагин
| S
| Аспарагин
| S
| Аспарагин
| S
|
| 6
| Цистеин
| ¾
| Цистеин
| ¾
| Цистеин
| ¾
|
| 7
| Пролин
|
| Пролин
|
| Пролин
|
|
| 8
| Лейцин
|
| Аргинин
|
| D-Аргинин
|
|
| 9
| Глюцинамид
|
| Глюцинамид
|
| Глюцинамид
|
|
РИС. 1-3.Химическое строение окситоцина, вазопрессина и 1 десмопрессина ацетата (ДА).
Выработка вазопрессина и окситоцина связана с синтезом специфических связывающих белков, называемых нейрофизинами. Одна молекула нейрофизина I связывается с одной молекулой окситоцина, а одна молекула нейрофизина II связывается с одной молекулой вазопрессина (Reeves et al, 1998). Комбинация нейрофизина и пептида, часто называемая нейросекреторным материалом, транспортируется вдоль аксонов гипоталамо-нейрогипофизарного нервного пути и сохраняется в гранулах нервных окончаний задней доли гипофиза (см. рис. 1-2). Высвобождение вазопрессина в кровь происходит после электрической активации крупноклеточных нейронов, содержащих АВП. Секреция происходит за счет экзоцитоза, при котором вазопрессин и нейрофизин II высвобождаются в кровь. В плазме комбинация нейрофизин-вазопрессин диссоциирует с высвобождением свободного вазопрессина. Почти все гормоны в плазме существуют в несвязанной форме, которая, благодаря своей относительно низкой молекулярной массе, легко проникает через периферические и клубочковые капилляры. Метаболическое расщепление АВП, по-видимому, опосредовано связыванием со специальными рецепторами АВП с последующим протеолитическим отщеплением пептида (Reeves et al, 1998). Выведение почками – второй способ удаления АВП из циркулирующей крови, составляющий примерно четверть от полного метаболического клиренса.
Дата добавления: 2016-09-20; просмотров: 1739;