Генетика поведения "простых" объектов
В случае очень многих мутаций экспрессия аномального гена прерывает эмбриогенез, и зародыш гибнет. Однако в ряде случаев оказывается возможным реконструировать функцию нормального алле-ля такого гена. Если удастся выделить мутации нескольких генов, экспрессирующихся на последовательных стадиях развития, то это может дать достаточно полную картину развития нервной системы. Подробная информация такого рода уже известна, она получена на интереснейшем генетическом объекте – микроскопической (2– 3 мм) почвенной нематоде Caenorhabditis elegans (Кайданов, Лучникова, 1981; Raiikin, Beck, 1992).
То, что удается узнать о генетическом контроле развития нервной системы и поведения, а также о генетическом контроле пластичности поведения у относительно простых объектов, делает, как мы надеемся, более понятной роль генотипа в формировании поведения насекомых и млекопитающих. Ниже приведены некоторые сведения по генетическому контролю физиологических реакций и "поведения" у бактерий, инфузорий и C.elegans.
 8.3.1. КИШЕЧНАЯ ПАЛОЧКА
8.3.1. КИШЕЧНАЯ ПАЛОЧКА
Разумеется, можно лишь весьма условно говорить о наличии "поведения" у бактерии. Очевидно, что движение бактерии как ответная реакция на градиент химических веществ – аттрактантов и репеллентов – является простейшим поведенческим актом, контролируемым генетически. Если жгутики – "органы движения" бактерии – вращаются в направлении "против часовой стрелки", они собираются в единый пучок, благодаря чему она совершает плавное поступательное движение. Если же направление вращения жгутиков сменяется на "по часовой стрелке", то пучок рассыпается, бактерия "спотыкается", останавливается и меняет направление движения.
Наличие рецепторов химических веществ позволяет бактериальной клетке, этому одноклеточному организму, получать информацию об их концентрации. В положительном градиенте привлекающих или отрицательном – неприятных веществ она движется достаточно плавно, практически не останавливаясь.
Таким образом, в репертуар "поведения" клетки-организма входят: возможность узнавания стимула, способность к обработке информации о его параметрах, формирование ответной реакции. Был описан целый ряд мутаций Escherichia со//, при которых перечисленные выше реакции нарушались. Условно их подразделяют на следующие категории:
а) нарушение рецепции (т.е. восприятия сигналов от аттрактантов или репеллентов); вещества, сходные по строению (сахара, ами нокислоты) действуют на свои специфические рецепторы, мута ции которых независимы друг от друга;
б) нарушение передачи сигнала от одного или более специфических рецепторов к жгутикам;
в) мутации, при которых жгутики способны к движению только в направлении "по часовой стрелке", что ведет к "спотыканиям"; реакции на химические вещества сохраняются, но при значи тельно более высоких концентрациях, чем в норме;
г) мутации, при которых жгутики способны к движению только "против часовой стрелки", и клетка, следовательно, может дви гаться только по прямой;
д) нарушение работы жгутиков или их способности к вращению. Аналитические исследования, проведенные в 70–80-е годы, пока зали, что на этом объекте можно исследовать наиболее примитив ные формы адаптации и фиксации предшествующего опыта, а также влияния условий окружающей среды на "поведение" клеток. По дробнее эти вопросы изложены Л. Эрман и П. Парсонсом (1984).
ИНФУЗОРИИ
Простейшие, и в частности инфузории, – это перспективный объект для исследования генетики наиболее примитивных реакций. Эти организмы крупнее и неизмеримо сложнее бактерий. В качестве генетического объекта обычно использовались инфузории рода Paramecium, в частности один из наиболее крупных видов – P.aureliwn.
Инфузории могут размножаться как бесполым (митотическим делением), так и половым путем. Половое размножение – конъюгация – обеспечивает генетическую изменчивость популяции, тогда как аутогамия – присущий простейшим способ самооплодотворения – позволяет получать полностью гомозиготных особей уже во втором поколении.
Движение инфузории осуществляется с помощью ресничек, покрывающих всю поверхность тела. Движение ресничек координируется общим физиологическим механизмом, связанным с возбудимой мембраной.
Мутации, выявленные у инфузории, влияют главным образом на характер движения. Особи "дикого типа" в норме перемещаются вперед короткими "проплывами", которые прерываются поворотами на 90*. Такие повороты рассматриваются как проявление спонтанных реакций избегания.
Генетические нарушения – мутации – выявляются именно в характере движений этих животных (рис. 8.5). При рецессивной мутации fast-2 инфузория очень быстро перемещается, совершая при этом большое число поворотов. Группа мутаций pawn (3 локуса, 62 мутации) характеризуется полным отсутствием реакции избегания, и клетка может двигаться только вперед (как пешка в шахматах). При paranoiac аномально усилены реакции избегания. Анализ физиологических процессов показал, что эти мутации нарушают нормальную функцию мембраны и почти не влияют на реснички. Существуют данные об аномальных электрофизиологических реакциях, соответствующих нарушениям разных типов.
Сложность строения этой группы организмов, в особенности инфузорий, у которых функции целого организма осуществляются в пределах одной клетки, не позволяет считать их удобным объектом для исследования мутаций, влияющих на поведение. Однако они могут быть использованы для оценки эффектов фармакологических агентов.
НЕМАТОДЫ
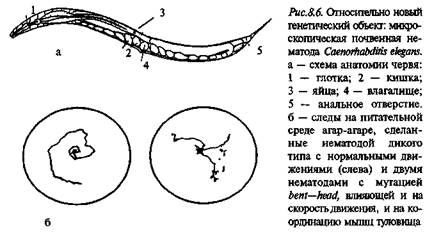
Свободно живущая почвенная нематода Caenorhabditis elegans была выбрана в качестве объекта исследования С. Бреннером в начале 70-х годов (рис. 8.6, а). Длина этого червя составляет около 1 мм, его репродуктивный цикл длится 3,5 суток. C.elegans – гермафродит, размножается самооплодотворением, откладывает до 300 яиц. Иногда в потомстве появляются особи только с мужскими половыми органами, которые спариваются с гермафродитами. Этот тип размножения дает дополнительные возможности для генетических исследований. У этого вида в гаплоидном наборе 5 аутосом и .^-хромосома. Построена почти полная карта генома C.elegans. По степени сложности строения тела и физиологических реакций эта нематода занимает промежуточное положение между Е. coli и дрозофилой. Геном C.elegans состоит из 8-Ю6 пар оснований, что в 20 раз больше, чем у E.coli, и примерно в 2 раза меньше, чем у дрозофилы. Около 83% генома нематоды составляют уникальные последовательности.
|
Для нематод характерна эвтелия, т.е. наличие в организме фиксированного числа клеток. Тело C.elegans состоит из 1000 клеток, причем 302 (по другим данным 350) из них – это нейроны. Генетическое, анатомическое и физиологическое исследование этого объекта, предпринятое С. Бреннером, имело несколько конкретных целей.
Во-первых, предполагалось создать пространственную картину связей всех клеток нервной системы с помощью электронномикро-скопического анализа многих тысяч серийных срезов.
Во-вторых, определялась химическая природа синапсов и выявлялись мутационные изменения, влияющие на поведение животного, последние сопоставлялись с изменениями в "рисунке" соединения клеток и/или химизме синапсов.
Описано несколько сот мутаций С. elegans, и примерно 2/3 из них затрагивают поведение. В спектр поведенческих изменений входят нарушения координации движений червя (рис. 8.6,6), аномалии строения тела, которые влекут за собой аномальные движения, изменения хемотаксиса, нарушение откладки яиц.
Примерно у половины мутантов с нарушенным поведением были найдены отклонения в строении нервной системы. Их можно разделить на 4 категории.
1. Изменения в клетках-предшественницах нейронов. Поскольку ход нормального развития нейронов из нейробластов изучен у этого вида достаточно хорошо, мутации в этих элементах легко обнару жить. Они могут проявляться в виде особенностей поведения на разных стадиях развития организма.
2. Изменения в специфичности синаптических связей. Поскольку соединения нейронов взрослой особи дикого типа уже известны, возникающие вследствие мутаций структурные изменения синапти ческих соединений можно с достаточной надежностью сопоставить с аномалиями поведения у таких животных.
3. Изменения в расположении отростков нейронов. Созданная полная топографическая карта нервной системы C.elegans позволяет выделить таких мутантов, у которых отростки нейронов идут к ано мальным мишеням. Такие мутации изменяют направление прораста ния и характер контактов у нейронов сразу нескольких классов. Оче видно, что изменения поведения при этом достаточно разнообразны.
4. Изменения в функции нейромедиаторов. Эту категорию эф фектов оказалось возможным проанализировать с развитием мето дов иммуноцитохимии и биохимии. Данные о медиа-горной специфич ности конкретных нейронов, как правило, оказываются неточными.
Электрофизиологические исследования этого объекта практически невозможны из-за его малого размера, поэтому сведения о нейрохимической специфичности нейронов того или иного класса сопоставляют с электрофизиологическими реакциями сходных групп клеток другой нематоды – Ascaris. У C.elegans обнаружены классические нейромедиаторы – ацетилхолин, ГАМК и биогенные амины.
Реакция на химические сигналы.Мутации, индуцированные химическими агентами, обычно бывают рецессивными и выявляются во втором поколении. Изучение поведения C.elegans и его изменений у мутантов показывает, что если у бактерий опознание химического вещества базируется на времени удержания следа, оставленного веществом на мембране и внутри клетки, то у C.elegans все обстоит сложнее. Так, например, мутант slow перемещается в 8 раз медленнее, чем особи дикого типа, но отыскание химических веществ, необходимых для питания, происходит у него без отклонений. Таким образом, для этой реакции время сохранения следа значения не имеет. Сопоставление градиента концентрации химического вещества вдоль тела червя также не может служить основой такой способности, так как мутанты, у которых изменены хеморецепторы хвостовой части тела, тем не менее, могут нормально отыскивать вещества. По всей видимости, основой реакции на химические вещества служит клинотаксис, т.е. движения, возникающие в результате оценки различий концентрации вещества с двух сторон тела. Поведенческим проявлением клинотаксиса, как считается, могут быть так называемые "рыскающие" движения головного конца.
Локомоция. Координация движений.В виду очень мелких размеров C.elegans основным методом выявления нарушений движения у него стала регистрация следов, которые червь оставляет на поверхности агара, служащего ему питательной средой. Примером мутаций, нарушающих движения, может служить roller, когда червь перемещается, переворачиваясь через головной и хвостовой концы, и bent head: при этой мутации изменена анатомия головного конца, и вместо обычных "рыскающих" движений перемещение идет по спирали. Движения червя обеспечиваются элементами брюшной нервной цепочки. Ее нейроны и их соединения хорошо описаны, известны также и эмбриональные клетки-предшественницы этих нейронов. Выделяют пять классов нейронов, которые подразделяются на два главных типа: возбудительные нейроны, инициирующие движение животного вперед или назад, и тормозные нейроны, координирующие сокращения мышц двух сторон тела.
Мутации группы Unc (uncoordinated) были одними из первых, описанными у C.elegans. Аномалии движений при неврологических мутациях очень перспективны для исследования организации движений в норме.
Фенотипически группа мутаций Unc подразделяется на 3 категории:
1) животные либо сохраняют неподвижность в покое, либо при сти муляции, которая в норме побуждает их двигаться вперед или назад, сокращаются все мышцы одновременно; такие системные мутации затрагивают функцию тормозных нейронов, в частности влияют на распределение их отростков (ипс-30); в других случаях нарушается синтез ГАМК;
2) животные не способны двигаться в одном из направлений (впе ред или назад), а при соответствующей стимуляции закручивают ся либо дорсальной, либо вентральной стороной кверху; установ лено, что в основе такого дефекта лежат аномальные связи ин тернейронов; в некоторых случаях отсутствует иннервация ряда мышц;
3) животные неспособны двигаться по прямой, не извиваясь; суще ствуют свидетельства того, что у таких мутантов аномалия затра гивает холинергические синапсы.
Реакция откладки яиц.Выход яиц из организма обеспечивают мышцы, иннервация которых идет от двух категорий нервных клеток, по-видимому, серотонинергических. Показано также, что ок-топамин эту реакцию тормозит, а его антагонист стимулирует. Описано около 40 мутаций, изменяющих процесс. При одной из них, например, нарушено собственно выведение яиц, и молодые особи вылупляются, оставаясь внутри родительского организма.
Мутации, влияющие на продолжительность жизни.Продолжительность жизни C.elegans – 18-21 день. Этот объект очень удобен для исследования молекулярно-генетических основ старения. Еще в начале работы с C.elegans была описана мутация age-1, при которой продолжительность жизни увеличивается до 34 дней.
Еще одна группа мутаций, влияющих на продолжительность жизни, связана с особенностями жизненного цикла этой нематоды. По окончании эмбриогенеза из яйца выходит личинка первой стадии (L1), которая затем проходит фазы L2, L3 и L4, после чего особь становится половозрелой. Однако в жизненный цикл этой нематоды может включаться еще одна, специфическая стадия – стадия покоящейся личинки, называемой дауером (dauer) – стадия d. В таком состоянии животное успешно переживает неблагоприятные средовые условия, например перенаселение или недостаток пищи. Стадия d может наступить в период после стадии L2 вместо L3. При восстановлении пригодных для жизни внешних условий из стадии d червь переходит в стадию L4.
В настоящее время предполагается, что в стадии d у животного происходит своеобразное выключение процессов старения, поскольку подобные покоящиеся личинки могут оставаться жизнеспособными на срок до 1 года. Выявлено семейство генов daf, участвующих в формировании стадии покоящейся личинки. Манипулируя условиями выращивания личинок с мутацией гена daf-2, можно получить особей, которые почти столь же жизнеспособны, как и особи дикого типа, но длительность их жизни значительно увеличена – до 43 дней. По существующим данным, у долгоживущих мутантов, отмечаются биохимические особенности, возможно связанные с изменением интенсивности процессов старения.
Пластичность поведения.Из всех описанных поведенческих реакций этой нематоды наиболее полно изучена реакция избегания в ответ на прикосновение. При стимуляции хвостового конца тела волоском червь реагирует продвижением вперед, при прикосновении к головному концу пятится назад. Неврологическим субстратом этих реакций является группа из 6 тактильных рецепторов, 5 пар интернейронов и 69 мотонейронов. Сочетание данных мутационного анализа и исследования фенотипа ряда мутантных форм (всего описано несколько сот таких мутаций) с микрохирургическими вмешательствами в ЦНСчервя с помощью лазерного луча позволило установить подробную схему соединений нервных элементов этой сети. Важно отметить, что эффект мутаций проявляется всегда в виде целого набора морфофункциональных изменений, и даже внимательный поиск не обнаруживает мутаций, эффект которых проявлялся бы в аномалии какого-то одного элемента этой сети.
Несмотря на относительную простоту, поведение C.elegans может видоизменяться в результате накопления индивидуального опыта. Экспериментально показано, что у этих животных могут происходить процессы: неассоциативного обучения – привыкание к регулярно действующему стимулу (механическое сотрясение), расторма-живание привыкания и сенситизация. Каждая из личиночных стадий имеет специфические особенности в формировании привыкания к тактильным стимулам. Обнаружение в будущем мутаций, влияющих на процессы неассоциативного обучения, в сочетании с беспримерным уровнем изученности морфофункциональных основ поведения этих животных даст новое понимание механизма пластичности реакций нервной системы (подробнее см.: Rankin, Best, 1992).
Относительно новый генетический объект, каким является C.elegans, очень удобен для изучения молекулярных основ патологии человека. В рамках большой исследовательской программы по генетическим механизмам болезни Альцгеймера были проведены эксперименты с получением трансгенных нематод. Как известно, одним из типичных проявлений патологии при болезни Альцгеймера являются отложения так называемого бета-амилоидного белка в виде бляшек.
У трансгенных особей C.elegans ген ипс-54, связанный с функцией мышц, был соединен с последовательностью оснований геном, кодирующим бета-амилоидный пептид. Этот чужеродный для нематоды ген экспрессировался, и его продукт был обнаружен. Анализ иммунореактивности в тканях таких генетически видоизмененных организмов позволил найти отложения, сходные с классическими бляшками бета-амилоидного белка, характерными для болезни Альцгеймера (см.: 8.6.5.5).
Таким образом, микроскопическая нематода как объект генетики поведения не только дает возможность исследовать нейроморфоло-гические основы примитивных реакций, но и становится удобным объектом для исследований по перенесению генетического материала (генной инженерии).
АПЛИЗИЯ
Аплизия, или морской заяц (Apfysia califomica), – объект, на котором проведены классические эксперименты по физиологии поведения моллюсков. В них продемонстрирована роль индивидуальных нервных клеток (так называемых индентифицированных нейронов) в формировании поведенческих реакций организма. Это новое направление – физиология поведения животных с "простыми нервными системами" (см.: Кенделл, 1980).
Исследование нейронных механизмов физиологических процессов у этого брюхоногого моллюска, например особенностей гормональной регуляции реакции откладки яиц, для которой известны все ее этапы, позволило перейти на молекулярно-генетический уровень и установить генную регуляцию группы конкретных ФКД из репертуара поведения этого животного.
Откладка яиц у аплизии обеспечивается цепью действий, включающую торможение локомоции, усиление дыхательных движений, специфические движения головы, с помощью которых жгут с яйцами вытягивается, сворачивается и приклеивается к субстрату. Эти события происходят вследствие активации сетей нейронов, которая является результатом координированного выделения нескольких пептидов. Пептиды, выделяющиеся железами репродуктивного тракта после копуляции, возбуждают нейросекреторные "пазушные" клетки. Они, в свою очередь, выделяют набор пептидов, которые воздействуют на разные нейроны моллюска. Возбуждение (или модуляция работы) пазушных клеток запускает сложную последовательность действий аплизии. Была выделена последовательность ДНК, кодирующая белок-предшественник, содержащий аминокислотные последовательности упомянутых пептидов. Считается установленным, что за формирование набора химических агентов, запускающих ФКД откладки яиц аплизии, отвечает один ген.
Сердечно-сосудистые реакции аплизии исследованы достаточно подробно в аспекте анализа функции пептидов (и их белков-предшественников), запускающих соответствующие нейроны. Пептид с последовательностью аминокислот Phe-Met-Arg-Phe, так называемый FMPF-иилш, описанный впервые у моллюсков, известен тем, что иммунореактивность к нему обнаруживается у всех представителей животного царства.
Генетика аплизии в классическом понимании не изучена: данные о генетической изменчивости особей, о выявленных мутациях и даже о строении ее генома пока отсутствуют. В то же время исследования, проведенные на этом моллюске, оказали мощное влияние на формирование современных нейрогенетических представлений не только в области генетики поведения дрозофилы, как наиболее генетически изученного экспериментального животного, но и для нейрогенетики в целом (подробнее см.: Nambu, Kirk, 1992).
8Л.
Дата добавления: 2016-08-07; просмотров: 1477;